 2020-05-12
2020-05-12 197
197
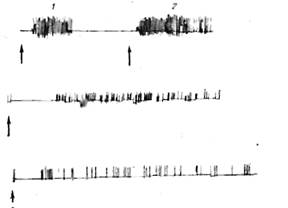
Рис. 23. Влияние различных доз сукцинилхолина на выраженность
двигательной реакции рыб под влиянием фенола (100 мг/л). 1,2 номера рыб.
Сверху вниз двигательная активность контрольных и подопытных карасей
при дозах сукцинилхолина 0,03, 0,0о и 0,6 мг/кг.
Стрелкой указан момент погружения карасей в раствор фенола
Вторая серия опытов поставлена на 42 карасях с флакседилом. Установлено, что только относительно высокие дозы препарата (от 2 до 6 мг/кг) вызывали торможение двигательной реакции рыб при погружении их в раствор фенола концентрацией 50-100 мг/л. Для купирования двигательной реакции карасей в растворе фенола (200-400 мг/л) необходимо было увеличить дозу флакесдила до 6-8 мг/кг.
Третья серия опытов с парамионом (15 рыб) показала, что дозы псепарата 3-6 мг/кг вызывают полное торможение двигательной реакции карасей в растворах фенола концентрацией 100 и 400 мг/л. Дозы парамиона 0,6, 1 и 2 мг/кг в ряде случаев также вызывали обездвижение рыб с потерей рефлекса равновесия, однако не тормозили развитие двигательной реакции карасей при погружении их в раствор фенола концентрацией 50-100 мг/л.
Анализируя данные в опытах с каждым из трех препаратов, вызывающих нервно-мышечную блокаду, можно сделать вывод, что в основе реакции рыб на остротоксичные концентрации фенола лежит не непосредственное раздражение фенолом мышц, тела, а рефлекторное движение под влиянием импульсов, исходящих из центральной нервной системы. Об этом свидетельствует снятие двигательной реакции рыб посредством блокирования передачи импульсов с нервов на мышцы в области мионеврального синапса. Из трех использованных нами курареподобных препаратов наиболее активным оказался сукцинилхолин из группы лептокураре, вызывающих стойкую деполяризацию концевой двигательной пластинки. По-видимому, сукцинилхолин выступает в качестве конкурента ацетилхолина в никотинчув-ствительных холинореактивных системах мышечных волокон, обусловливая более эффективное функциональное разобщение нервной и мышечной систем, чем это отмечено в опытах с флакседилом или парамионом. Результаты этих опытов свидетельствуют также о важной роли ацетилхолинового метаболизма в механизмах развития внешнего симптомокомплекса фенольной интоксикации рыб.
Хеморецепторы
Согласно современным представлениям сенсорной физиологии первичное восприятие химических раздражителей осуществляется специализированными хеморецепторами — специальными образованиями, в которых происходит трансформация энергии раздражающего стимула в сигналы, несущие нервным центрам информацию о действующем агенте. У рыб, как и у других позвоночных животных, восприятие многочисленных химических раздражителей, поступающих в воду, осуществляется хеморецепторами. Афферентная импульсация, возникающая в хеморецепторах, направляется в центральную нервную систему, в результате чего организм получает информацию о присутствии в водной среде тех или иных химических соединений.
Химическая рецепция рыб включает обоняние, вкус и общее химическое чувство [205а]. Разделение первых двух видов химической рецепции рыб экспериментально обосновано с морфологических и физиологических позиций и потому общепризнано. Что касается выделения общего химического чувства, отличного от вкусового, то его правомерность все еще дискутируется [2, 37].
Важнейшим физиологическим свойством любого рецептора является чувствительность к адекватному стимулу. Уровень чувствительности оценивается порогом возбуждения, т. е. минимальной величиной стимула, способной вызвать возбуждение. Величина порога обратна величине возбудимости. Существуют различные методы регистрации обонятельных порогов пахучих веществ (условные рефлексы, поведенческие реакции, электрическая активность). С их помощью удалось установить, что рыбы обладают исключительно высокой чувствительностью обонятельных рецепторов, в сотни раз превышающей чувствительность обонятельной рецепции человека [205а]. Это позволяет рыбам воспринимать чрезвычайно малые концентрации химических веществ, обладающих запахами.
Уже в первоначальных работах [585, 586] с помощью метода условных рефлексов выявлена способность рыб (тупоносые хибо-ринхусы) обнаруживать фенол концентрацией 0,01 мг/л. Некоторые особи после длительной тренировки отличали фенол от парахлорфенола при концентрации этих веществ 0,0005 мг/л, что значительно ниже порога чувствительности для человека.
Гольяны различают запахи эвгенола и фенилэтилового спирта концентрацией 6•10-14 и 4,3•10-14 мг/л (соответственно), т.е. концентрацией, в 250 раз более низкой, чем человек [726]. Дальневосточные лососи обнаруживают морфолин концентрацией 1•10-12 мг/л [583]. Еще более высокая чувствительность рыб выявлена в опытах с α-ионопом (0,2•10-15 М) и β-фенилэтилалкоголем (0,5•10-18мг/л). Угри воспринимали β-фенилэтилалкоголь даже концентрацией 3•10-20 М, т. е. когда в ольфакторном мешке могли находиться лишь 2-3 молекулы этого вещества [815]. Чрезвычайно высока чувствительность плотвы к бензолу и особенно его производным [691]. Рыбы воспринимали 1, 3, 5-тринитробензол концентрацией 1,4•10-16 М, 1,3-динитробензол — 7•10-16 М, нитробензол — 7,3•10-6 M, бензол — концентрацией 20-10-6 М. Несколько ниже, но достаточно высокая чувствительность этого вида рыб к фенолу (9,4 • 10-6 М) и его производным — резорцину (54,2 • 10-6 М) и флороглюцину (76•10-6 М). Караси и данио (Danio malabaricus) обнаруживают кумарин даже при таких низких концентрациях, как 1•10-17 — 1•10-18 М, оказывающих на них репеллентное действие [593]. Любопытно, что более высокие концентрации этого вещества (от 10-12 до 10-2 М) привлекают рыб.
Электрофизиологическими исследованиями подтверждена высокая чувствительность обонятельной рецепции рыб, которые хорошо воспринимают различные органические соединения — спирты, кетоны, альдегиды, кислоты, эфиры, фенол и его производные [205а]. Так, с помощью электрофизиологичеокого метода показано, что бутиловый спирт (концентрацией 10-15 М), уксусная кислота (10-13 М), морфолин (10-11 М) вызывают четко выраженные реакции в обонятельной системе сомов [465]. По мнению Дж. Будро [465], истинные пороги лежат ниже этих величин, и есть основания согласиться с этим мнением. Известно, что при электрофизиологических исследованиях периферических отделов обонятельного анализатора пороговые интенсивности обонятельного раздражения, вызывающего ответ, обычно значительно превосходят порог, измеренный по поведенческим реакциям, что объясняется скорее всего несовершенством техники регистрации.
Необонятельная химическая рецепция рыб (вкус и общая химическая чувствительность) так же высока, как и обонятельная. Рецепторный аппарат вкусовой системы рыб представлен вкусовыми почками, расположенными в слизистой оболочке ротовой полости, на жабрах и наружной поверхности тела. Кроме того, восприятие химических раздражителей осуществляется свободными нервными окончаниями тройничного, блуждающего и спинномозговых нервов. Основными проводниками химической необонятельной рецепции служат блуждающий, языкоглоточный и лицевой нервы [37]. Наружные вкусовые почки рыб высокочувствительны к различным веществам. Например, вкусовая чувствительность у гольянов по отношению к глюкозе в 1,6 раза, к фруктозе в 2,5 раза, хинину в 24 раза, а к поваренной соли в 205 раз выше, чем у человека [563].
Электрофизиологическими исследованиями показаны способность пресноводных рыб воспринимать химические вещества в очень низких концентрациях [647] и наличие в небном органе карпа особой хеморецепторной системы, отвечающей специфично на разведенные растворы электролитов [646], интенсивность ответа зависит от валентности аниона. В опытах на пресноводном сомике выявлено существование двух типов кислотных волокон, одни из которых отвечают только на органические кислоты, а другие — на соляную [813, 814]. Изучение электрической активности лицевого и небного нервов атлантического лосося при химическом раздражении области рыла и небного органа показало, что под влиянием слабых растворов солей, минеральных и органических кислот в афферентных нервах возникают разряды, величина которых зависит от концентрации и химической природы вещества [806].
Многолетнее изучение химической необонятельной рецепции рыб, в частности ее центральных механизмов с помощью электрофизиологических методов, и особенностей анализа различных химических веществ при использовании метода условных рефлексов ведется сотрудниками кафедры физиологии высшей нервной деятельности Ленинградского университета [2, 37]. Анализируя результаты этого направления исследований, Н. Е. Василевская [37] приходит к вполне обоснованному выводу «о высокой специализации химического анализатора, обеспечивающей распознавание очень небольших концентраций химических веществ, воздействующих на кожную поверхность» (с. 52).
Имеющиеся экспериментальные данные однозначно свидетельствуют о чрезвычайно высокой чувствительности рыб к различным группам химических веществ, что ставит их в особое положение по отношению к другим гндробионтам, в частности к водным беспозвоночным. Конечно, последние также обладают определенным уровнем чувствительности к токсикантам, поскольку химическая раздражимости— общее свойство живых клеток. Однако среди гидробионтов только рыбы обладают высоко развитой центральной нервной системой и специализированными анализаторными системами, в частности химическим анализатором, специальное назначение которого — информировать организм о присутствии в окружающей среде разнообразных химических веществ.
Возникшие в процессе эволюции хеморецепторы приобрели особую чувствительность и специфичность в восприятии отдельных свойств химических веществ (вкус, запах), что позволяет им тонко анализировать и своевременно реагировать на химические изменения в окружающей водной среде. Вполне понятно, что чем быстрее и точнее организм получает информацию о состоянии внешней среды и ее изменении, тем быстрее он может включить защитные и компенсаторные механизмы в ответ на эти изменения, тем выше его возможности выживания в экстремальных условиях.
Предпринятый нами экспериментальный анализ роли нервной системы в реакции рыб на фенольное раздражение позволил выявить центральное звено и эфферентную часть рефлекторной дуги этой реакции. Естественно, возникал вопрос, какова афферентная часть, т. е. чувствительные пути, по которым передаются импульсы в центральную нервную систему, и где локализованы хеморецепторы, воспринимающие фенольное раздражение.
Как мы уже отмечали, восприятие химических раздражителей осуществляется у рыб рецепторными аппаратами обонятельной и вкусовой систем, а также свободными нервными окончаниями тройничного, блуждающего и спинномозговых нервов. Можно было предположить, что вкусовые почки и рецепторы общего химического чувства, предназначенные для анализа химического состава или «вкуса» воды, расположенные по всему телу и на пути прохождения воды для дыхания, будут играть ведущую роль в восприятии фенольного раздражения. Вкусовые почки, расположенные на поверхности тела, иппервируются лицевым нервом, а в тех из них, которые находятся в полости рта, глотки и жаберного аппарата, оканчиваются чувствительные волокна языко, глоточного и блуждающего нервов. Все это давало основание надеяться; что выключение периферического звена вкусового анализатора может оказать существенное влияние на скорость и время проявления феиолыюй реакции.
Для проверки этого предположения мы выполнили опыты по фармакологическому выключению рецепторов поверхности тела, а также чувствительных нервных окончаний, расположенных в полости рта, глотки и жаберного аппарата. Использовали кокаин, оказывающий глубокий местноанестезирующий эффект и вызывающий полную потерю чувствительности рецепторов в течение нескольких минут. В опытах, проведенных нами па 23 карасях, применяли 0,01- и 0,1%-ный растворы кокаина при экспозициях 5 и 15 мин (поверхностные рецепторы) и 3 и 5 мин (чувствительные нервные окончания в полости рта и жаберного аппарата). Ни в одном случае нам не удалось предупредить или снять фенольную локомоторную реакцию. Двигательная активность кокаинизированных карасей развивалась так же быстро и с такой же интенсивностью, как и у контрольных особей.
Не произошло существенных изменений в реакции рыб на фенол и после двусторонней перерезки или удаления п. vagi-n. lateralis ипнервирующего боковую линию туловища. Перерезка латеральной ветви блуждающего нерва производилась вблизи от его выхода из черепа. По данным 10 опытов, локомоторная реакция у подопытных особей развивалась под влиянием фенола в полной мере и была идентична реакции интактных карасей. Единственное незначительное различие касалось латентного периода наступления двигательной реакции, который у оперированных карасей оказался на 17,5% больше, чем у контрольных (114 и 97 соответственно).
Поскольку нами достоверно установлена рефлекторная природа фенольной реакции рыб, выявлены центральное звено и эфферентные пути рефлекторной дуги, можно предположить, что либо фенольное раздражение воспринимается хеморецепторами, локализованными не на поверхности тела, а в более глубоко расположенных тканях, либо фенол действует как гуморальный автоматический раздражитель непосредственно на центральную нервную систему.








