2020-05-12
2020-05-12 200
200
Выявленные методом условных рефлексов изменения функционального состояния центральной нервной системы под влиянием различных токсикантов были подтверждены с помощью электрофизиологическйх методов исследования, в основе которых лежит регистрация фоновой электроэнцефалограммы (ЭЭГ) и вызванных потенциалов (ВП). Одна из первых работ в этом плане выполнена на золотых рыбках, отравленных ДДТ [430]. У рыб, помещенных в токсический раствор, отмечено увеличение амплитуды и снижение преобладающей частоты ЭЭГ мозжечка во время нарушения рефлекса равновесия. После помещения рыб в чистую воду биоэлектрическая активность мозжечка и рефлекс равновесия восстанавливались в течение длительного периода времени (от 1 до 5 сут), определяемого предшествующей экспозицией рыб в токсическом растворе ДДТ. Электрофизиологический метод оценки функционального состояния различных отделов центральной нервной системы рыб находит все более широкое применение при изучении степени токсичности пестицидов [424, 431, 742, 197, 252], нефти [196, 197, 252, 114],сточных вод сложного состава [242].
Еще в конце 50-х годов было обнаружено понижение чувствительности зрительного восприятия у лосося (Oncorhynehus kisutch), отравленного ДДТ [416]. В дальнейшем с помощью метода условных рефлексов удалось подтвердить влияние ДДТ на зрение рыб [704].
Скармливание радужной форели 30, 50 и 80% токсической дозы ДДТ вместе с кормом приводило через 3 сут к потере способности различать яркий и тусклый свет. Электрофизиологический анализ механизма действия фосфорорганических пестицидов (карбофоса и хлорофоса) на зрение рыб (карп, ротан) показал, что их влияние проявляется на всех уровнях зрительного анализатора [197, 252]. Выраженность токсического эффекта определяется концентрацией, временем действия токсиканта и исходным функциональным состоянием подопытных рыб.
Сходные по направленности и глубине изменения функционального состояния мозга, в частности центральных отделов зрительного анализатора, обнаружены С. И. Никоноровым и А. С. Лукьяновым [252] у морских рыб (мерланг, колючая акула) под влиянием водорастворимых фракций нефти. В качестве показателя функционального состояния центральной нервной системы были избраны вызванные потенциалы переднего и крыши среднего мозга в ответ на электрическую стимуляцию зрительного нерва и адекватную стимуляцию глаза рыб светом. У отравленных рыб отмечены достоверное увеличение амплитуды вызванного потенциала переднего и крыши среднего мозга в ответ на электрическую стимуляцию зрительного нерва (рис. 25) и менее выраженные изменения латентных периодов максимумов негативного и позитивного компонентов вызванных потенциалов переднего мозга.
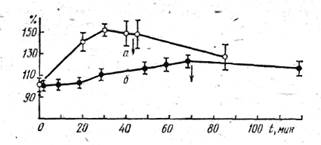
Рис. 25. Изменения амплитуды вызванного потенциала переднего (а)
и крыши среднего мозга (б) в процессе воздействия водорастворимых фракций нефти (5 мг/л)
на центральную нервную систему черноморской колючей акулы.
Стрелками отмечено введение и выведение токсического вещества из системы [252]
Обнаружены также изменения амплитуды и компонентного состава вызванных потенциалов крыши среднего мозга (рис. 26), отражающие процессы переработки информации в периферическом отделе зрительной системы рыб — сетчатке глаза. Таким образом, как и в опытах с пестицидами, проведенными на пресноводных рыбах, у морских рыб под влиянием токсического воздействия нефти происходят функциональные сдвиги на всех уровнях зрительного анализатора, и наиболее выражены они в структурах переднего мозга 1252].
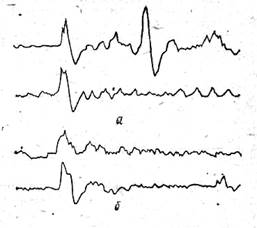
Рис. 26. Изменение вызванного потенциала крыши среднего мозга в ответ
на адекватную стимуляцию глаза светом при воздействии
водорастворимых фракций нефти (5 мг/л) на центральную нервную систему
черноморской колючей акулы: в — вызванный потенциал на включение света:
б— то же, на выключение света; нижние записи вызванных потенциалов (а, б)
зарегистрированы после воздействия (калибровка времени 200 мс, амплитуды — 500 мкВ).
Отклонение потенциала вверх соответствует негативности [252]
Обстоятельное исследование характера влияния водорастворимых фракций нефти в субтоксических концентрациях (0,05 — 0,12 мг/л) на функциональную деятельность зрительного анализатора у проходных (белуга и осетр) и пресноводных (сазан) рыб выполнено Р. Ю. Касимовым и сотрудниками [114]. Они также регистрировали вызванные потенциалы в крыше среднего мозга при раздражении зрительного нерва у отравленных нефтью рыб. Концентрация нефти свыше 0,05 мг/л вызывала у молоди белуги уже на 1-е сутки резкое снижение амплитуды длительности и периода пика вызванного потенциала, а также увеличение латентного периода. При возрастании времени контакта рыб с токсикантом до 2 сут отмечены еще более значительные нарушения конфигурации вызванного потенциала крыши среднего мозга. Восстановление основных параметров вызванного потенциала отмечено только на 6-7-е сутки после перевода рыб в чистую воду. Авторы пришли к выводу, что под воздействием даже небольших концентраций нефтяного загрязнения происходит как бы «притупление» анализаторной системы, вследствие чего реакция рыб на различные раздражители таожет значительно меняться.
Несколько ранее сходные данные были получены и при электрофизиологическом изучении функционального состояния обонятельного анализатора. Так, в опытах на препарате головы миксины с помощью электроэнцефалограммы установлено, что длительное воздействие на обонятельные рецепторы раствором хлористого калия (1•10-6 М) снижает их ответ на пахучие раздражители [533]. Этим же методом показано, что 1%-ный раствор морфолина блокирует обонятельную рецепцию рыб к действию пахучих химических раздражителей [580]. Вполне понятно, что направленность изменений функционального состояния центральных и периферических отделов того или иного анализатора рыб под влиянием разных токсикантов может быть различной. Например, ДДТ повышает чувствительность терморецепторов боковой линии форели, которая, по мнению авторов [742], имеет центральное происхождение.
Многие токсиканты, поступающие в рыбохозяйственные водоемы, уже в субтоксических концентрациях могут изменять функциональное состояние центральной нервной системы в целом и отдельных анализаторных систем, блокировать или выводить из строя периферические отделы этих анализаторных систем, что будет неизбежно отражаться на поведенческих реакциях рыб, обусловленных абиотическими и биотическими факторами водной среды.
Сегодня имеется уже достаточно фактов, подтверждающих это. Прежде всего привлекают внимание данные, согласно которым у рыб под влиянием различных токсикантов (пестициды-ДДТ, эльдрин, полихлориро-ваниые бифенилы; фенол) происходит изменение температурного предпочтения (729, 824, 479, 743, 712, 730, 744].
Опытами, выполненными на атлантическом лососе, выявлены разнонаправленные изменения термопреферендума после выдерживания их в растворах ДДТ различных концентраций [729]. После 24-часового контакта с низкими концентрациями пестицида (0,005-0,01 мг/л) лососи предпочитали температуру на 3-5° С ниже, чем контрольные, а после контакта с более высокими концентрациями (0,02-0,05 мг/л), напротив, — на 11-13° С выше, чем контрольные. Результаты этих наблюдений были подтверждены в опытах на атлантическом лососе и гольце [824], но при других концентрациях ДДТ. После выдерживания в течение 1 сут в растворе ДДТ концентрацией 2-10 мг/л рыбы предпочитали более низкую температуру, а после воздействия на них высоких концентраций ДДТ (до 200 мг/л) — повышенную температуру. По данным Р. Петерсона [743], эльдрин вызывает у атлантического лосося и ручьевой форели сдвиг температурного предпочтения в зону пониженных (на 3°С) температур, а ДДТ и его производные, напротив, — в зону повышенных (на 4-5° С) температур. Автор высказывает предположение о том [744], что изменения температурного предпочтения у рыб, подвергшихся токсическому воздействию, носят в известной мере адаптивный характер: изменение термопреферендума в сторону пониженных температур происходит под влиянием веществ, токсичность которых увеличивается при высоких температурах. Однако имеются данные [479], согласно которым под влиянием ДДТ и его метаболитов у рыб вообще нарушается способность выбора температур в течение довольно длительного времени (до 9 сут).
Нарушение различных сторон анализаторной деятельности центральной нервной системы рыб, их способности воспринимать и адекватно реагировать на изменения окружающей среды под влиянием субтоксических концентраций химических загрязнений может оказать существенное влияние на сложные формы поведения рыб в естественных водоемах (нагульные, зимовальные и нерестовые миграции, ориентацию, взаимоотношение с хищными видами рыб, обнаружение и избегание токсических веществ) [163]..