 2020-05-25
2020-05-25 228
228нант функционального резерва сердца по его специфической функции — перемещению крови в системе. При увеличении резервного объема, соответственно, увеличивается максимальный систолический объем, который может быть выброшен из сердца в условиях его интенсивной деятельности.
Регуляторные влияния на сердце реализуются в изменении систолического объема путем воздействия на сократительную силу миокарда. При уменьшении мощности сердечного сокращения систолический объем снижается.
У человека при горизонтальном положении тела в условиях покоя систолический объем составляет от 60 до 90 мл (табл. 9.3).
Таблица 9.3. Некоторые параметры системной гемодинамики и насосной функции сердца у человека (в условиях основного обмена)
| Сердечный выброс, л/мин | 4,0- | -6,0 |
| Частота сердечных сокращений, мин-1 | 60- | -80 |
| Конечно-диастолический объем, мл | 65- | -130 |
| Конечно-систолический объем, мл | 40- | -60 |
| Ударный объем сердца, мл | 60- | -90 |
| Давление, мм рт. ст. | Аорта | Легочный ствол |
| систолическое | 110—120 | 25-30 |
| диастолическое | 60-75 | 6-12 |
| среднее | 70-80 | 9-17 |
| Общее периферическое сопротивление сосу | Большой круг | Малый круг |
| дов, дин • с • см-5 | 1200-1700 | 150-350 |
| Давление, мм рт. ст. | Левое предсердие | Правое предсердие |
| конечно-с истол ическое | 10-12 | 3-7 |
| конечно-диастолическое | 0-6 | 0-2 |
| Давление, мм рт. ст. | Левый желудочек | Правый желудочек |
| конечно-систолическое | 110-120 | 25-30 |
| конечно-диастолическое | 9-12 | 6-10 |
| Максимальная скорость прироста давления | ||
| (dP/dt,nax), мм рт. ст./с | 2000-2500 | 350-400 |
| Работа за систолу, Дж | 0,9-1,0 | 0,2 |
9.1.3.4. Частота сердечных сокращений (пульс)
Частота сердечных сокращений (пульс) в покое составляет от 60 до 80 ударов в минуту. Регуляторные влияния, вызывающие изменения частоты сердечных сокращений, называются хронотропными, а изменения силы сокращений сердца — инотропными.
Повышение частоты сердечных сокращений является важным адаптационным механизмом увеличения МОК, осуществляющим быстрое приспособление его величины к требованиям организма. При некоторых экстремальных воздействиях на организм сердечный ритм может повышаться в 3—3,5 раза по отношению к исходному.
Важным показателем системной гемодинамики и сердечной деятельности является работа сердца за систолу.
9.1.3.5. Работа сердца
Миокард желудочков в период изометрического сокращения сообщает находящейся в них крови потенциальную энергию, которая в период изгнания крови переходит в кинетическую энергию движущейся крови. Большая часть последней переходит в сосудистой системе в потенциальную энергию растянутых эластических стенок аорты и легочной артерии, и лишь небольшое количество ее (2—5 %) сохраняется в виде кинетической энергии движущейся крови.
Поскольку вся полезная работа сердца переходит в энергию крови, полную работу сердца за систолу можно рассчитать как сумму потенциальной и кинетической энергии крови. Потенциальная часть работы сердца вычисляется как произведение ударного объема сердца на среднее артериальное давление. Кинетическая часть работы сердца рассчитывается как половина произведения массы выбрасываемой из сердца крови за систолу на квадрат средней скорости изгнания крови из сердца. В покое кинетическая часть работы сердца составляет лишь 2—5 % от полной работы сердца, поэтому ее обычно не учитывают при расчетах. Однако при физических нагрузках кинетическая часть работы сердца у человека может составлять до 30 % от полной работы сердца за систолу. Рассчитанная таким образом работа характеризует деятельность левой половины сердца. Работа правых отделов сердца составляет около 25 % от этой величины.
9.1.3.6. Сократимость
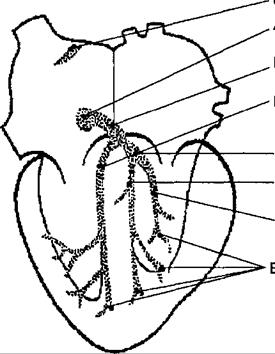
Сократимость, т. е. способность сокращаться, характерная для всех разновидностей мышечной ткани, реализуется в миокарде благодаря трем специфическим свойствам сердечной мышцы: автоматизм — способность клеток водителей ритма генерировать импульсы без каких-либо внешних воздействий; проводимость — способность элементов проводящей системы к электротонической передаче возбуждения; возбудимость — способность кардиомиоцитов возбуждаться в естественных условиях под влиянием импульсов, передаваемых по волокнам Пуркинье (рис. 9.5). Важной особенностью возбудимости сердечной мышцы является длительный рефракторный период (полное исчезновение или резкое снижение возбудимости кардиомиоцитов после их предыдущего сокращения), гарантирующий ритмический характер последующего сокращения.
9.1.3.6.1. Автоматизм и проводимость миокарда
В области правого предсердия, а также на границе предсердий и желудочков располагаются участки, ответственные за возбуждение сердечной мышцы. Автоматизм сердца имеет миогенную природу и обусловлен спонтанной активностью части клеток его атипической ткани.
Указанные клетки образуют скопления в определенных участках миокарда. Наиболее важным в функциональном отношении из них является синусный, или синоатриальный, узел, расположенный между местом впадения верхней полой вены и ушком правого предсердия. В нижней части межпредсердной перегородки, непосредственно над местом прикрепления септальной створки трехстворчатого клапана, располагается атриовентрикулярный узел. От него отходит пучок атипических мышечных волокон, который пронизывает фиброзную перегородку между предсердиями и переходит в узкий длинный мышечный тяж, заключенный в межжелудочковую перегородку. Он называется атриовентрикулярным пучком, или пучком Гиса. Пучок Гиса разветвляется, образуя две ножки, от которых приблизительно на уровне середины перегородки отходят волокна Пуркинье, также образованные атипической тканью и формирующие субэндокардиальную сеть в стенках обоих желудочков (см. рис. 9.5).
Функция проводимости в сердце имеет электротоническую природу. Она обеспечивается низким электрическим сопротивлением щелевидных контактов (нексусов) между элементами атипического и рабочего миокарда, а также в области вставочных пластинок, разделяющих кардиомиоциты. В результате сверхпороговое раздражение любого участка вызывает генерализованное возбуждение всего миокарда. Это позволяет считать ткань сердечной мышцы, морфологически разделенную на отдельные клетки, функциональным синцитием.
Возбуждение миокарда зарождается в синоатриальном узле, который называют водителем ритма, или пейсмекером первого порядка, и далее
|
|
 |  |
распространяется на мускулатуру предсердий с последующим возбуждением атриовентрикулярного узла, который является водителем ритма второго порядка. Скорость распространения возбуждения в предсердиях составляет 1 м/с. При переходе возбуждения на атриовентрикулярный узел имеет место так называемая атриовентрикулярная задержка, составляющая 0,04— 0,06 с. Механизм атриовентрикулярной задержки состоит в том, что проводящие ткани синоатриального и атриовентрикулярного узлов контактируют не непосредственно, а через волокна рабочего миокарда, для которых характерна более низкая скорость проведения возбуждения. Последнее распространяется далее по ножкам пучка Гиса и волокнам Пуркинье, передаваясь на мускулатуру желудочков, которую оно охватывает со скоростью 0,75—4,0 м/с. В силу особенностей расположения волокон Пуркинье возбуждение сосочковых мышц происходит несколько раньше, чем оно охватывает стенки желудочков. Благодаря этому нити, удерживающие трехстворчатый и митральный клапаны, оказываются натянутыми раньше, чем на них начинает действовать сила сокращения желудочков. По той же причине наружная часть стенки желудочков у верхушки сердца возбуждается несколько раньше участков стенки, прилежащих к ее основанию. Таким образом, волна возбуждения последовательно охватывает различные отделы сердца в направлении от правого предсердия к верхушке. Однако указанные сдвиги во времени крайне невелики и обычно принимается, что весь миокард желудочков охватывается возбуждением одновременно.
9.1.3.6.2. Мембранная природа автоматии сердца
Возбудимость клеток проводящей системы и рабочего миокарда имеет ту же биоэлектрическую природу, что и в поперечно-полосатых мышцах. Наличие заряда на мембране здесь также обеспечивается разностью концентраций ионов калия и натрия возле ее внешней и внутренней поверхности и избирательной проницаемостью мембраны для этих ионов. В покое мембрана кардиомиоцитов проницаема для ионов калия и почти непроницаема для ионов натрия. В результате диффузии ионы калия выходят из клетки и создают положительный заряд на ее поверхности. Внутренняя сторона мембраны становится электроотрицательной по отношению к наружной.
В клетках атипического миокарда, обладающих автоматией, мембранный потенциал способен спонтанно уменьшаться до критического уровня, что приводит к генерации потенциала действия. В норме ритм сердечных сокращений задается всего несколькими наиболее возбудимыми клетками синоатриального узла, которые называются истинными водителями ритма, или пейсмекерными клетками. В этих клетках во время диастолы мембранный потенциал, достигнув максимального значения, соответствующего величине потенциала покоя (60—70 мВ), начинает постепенно снижаться. Этот процесс называют медленной спонтанной диастолической деполяризацией. Она продолжается до того момента, когда мембранный потенциал достигает критического уровня (40—50 мВ), после чего возникает потенциал действия.
Для потенциала действия пейсмекерных клеток синоатриального узла характерны малая крутизна подъема, отсутствие фазы ранней быстрой реполяризации, а также слабая выраженность «овершута» и фазы «плато». Медленная реполяризация плавно сменяется быстрой. Во время этой фазы мембранный потенциал достигает максимальной величины, после чего вновь возникает фаза медленной спонтанной деполяризации (рис. 9.6).









