 2020-05-25
2020-05-25 604
604В мышечных клетках человека, как и в любых других видах клеток, происходит энергетический обмен, при котором богатые энергией питательные вещества усваиваются и химически преобразуются, а конечные продукты обмена с более низким содержанием энергии выделяются из клетки. Высвобождающаяся при этом энергия в мышцах обеспечивает их сокращение. Основным источником энергии для мышечного сокращения при физической работе является гидролиз АТФ. Макроэргические фосфатные связи молекулы АТФ нестойкие и концевые фосфатные группы легко отщепляются, при этом освобождается энергия (7—10 ккал/моль АТФ). Присоединение молекул АТФ к головкам миозиновых мостиков и последующий гидролиз АТФ являются ключевым моментом в процессе преобразования химической энергии фосфатных связей в механическую энергию мышечного сокращения. В нормальных физиологических условиях мышечная активность как низкой, так и высокой мощности не сопровождается истощением запасов энергии макроэргических соединений. Это связано с процессами ресинтеза АТФ. Различают аэробный и анаэробный пути ресинтеза АТФ.
22.1.1. Анаэробный путь ресинтеза АТФ
При кратковременной физической работе большой мощности ресинтез АТФ происходит анаэробным путем, в котором выделяют три процесса: фосфогенный, миозинкиназный и гликолиз.
Фосфогенный анаэробный процесс (креатинфосфокиназная реакция). В мышцах наряду с АТФ содержится другое макроэргическое фосфорное соединение — креатинфосфат, являющееся вторым после АТФ по значимости макроэргическим фосфатным соединением. Креатинфосфат адсорбируется на сократительных белках миофибрилл и на мембране саркоплазматического ретикулума. С тонким филаментом — актином связан фермент креатинфосфокиназа, который катализирует реакцию перефосфори- лирования между креатинфосфатом и аденозидифосфатом (АДФ), образующимся при гидролизе АТФ.
Креатинфосфат + АДФ--------------------------- > АТФ + Креатин
Креатинфосфокиназная реакция первой включается в процесс ресинтеза АТФ в момент начала мышечной работы и протекает с максимальной скоростью до тех пор, пока запасы креатинфосфата в мышцах не будут исчерпаны примерно на 30 % (в течение 5—6 с). Затем скорость креатинфосфокиназной реакции начинает уменьшаться и в процесс ресинтеза АТФ вступают гликолиз и аэробный механизм.
Фосфогенный аэробный процесс ресинтеза АТФ играет решающую роль в энергетическом обеспечении кратковременной физической работы максимальной мощности (например, подъем и перенос тяжелых грузов), в процессе быстрого перехода от покоя к работе, во время финишного ускорения у спортсменов.
Анаэробный гликолиз. Как только в процессе мышечной работы креатинфосфокиназная реакция перестает обеспечивать необходимую скорость ресинтеза АТФ и в саркоплазме мышечных волокон увеличивается концентрация молекул АДФ, происходит активация ресинтеза АТФ за счет анаэробного гликолиза. Источником энергии для ресинтеза АТФ являются запасы гликогена в миоцитах и глюкоза плазмы крови. Повышение концентрации АДФ в саркоплазме, а также увеличение содержания ионов Са++ активируют ферменты фосфорилазу и гексокиназу. Молекулы этих ферментов локализованы на мембране саркоплазматического ретикулума, а также на других внутриклеточных компонентах.
В процессе гликолиза образуется до 2,5 кДж/кг/мин. Гликолиз как источник энергии мышечного сокращения эффективно обеспечивает сократительный процесс с 20-30-й секунды после начала сокращения и способен обеспечивать мышечную активность высокой мощности до 2—2,5 мин. Скорость гликолиза угнетается под влиянием повышения концентрации молочной кислоты в миоцитах и уменьшения pH в саркоплазме. Накопление молочной кислоты при анаэробной работе находится в прямой зависимости от мощности и общей продолжительности физической работы.
Увеличение количества молочной кислоты в саркоплазматическом пространстве мышц вызывает рост осмотического давления в саркоплазме, при этом вода из межклеточной среды поступает внутрь мышечных волокон и вызывает их набухание. Значительные изменения осмотического давления в мышцах могут быть причиной болевых ощущений.
Миокиназная реакция. Физическая активность высокой мощности, а также развитие утомления (фаза истощения) приводят к повышению в саркоплазме концентрации молекул АДФ, которые являются источником ресинтеза АТФ:
Аденилатциклаза
АДФ + АДФ--------------------------- > АТФ + АМФ.
Миокиназная реакция, как и креатинфосфокиназная, легко обратима и используется в организме для уменьшения резких перепадов в скорости образования и использования АТФ. В случае появления в клетке избытка АТФ он быстро устраняется с помощью миокиназной реакции.
22.1.2. Аэробный гликолиз
Примерно 90 % запасов АТФ в миоцитах образуется в процессе аэробного ресинтеза, т. е. при участии кислорода. Ферментные системы аэробного обмена расположены в основном в митохондриях клеток. К числу субстратных циклов окисления относятся: гликолитическое расщепление углеводов, заканчивающееся образованием пировиноградной кислоты, окислительное декарбоксилирование пировиноградной кислоты, цикл превращений трикарбоновых кислот, окислительное дезаминирование аминокислот, [J-окисление жирных кислот. Суть химических превращений в субстратных циклах заключается в постепенном преобразовании исходного субстрата в форму, доступную действию специфических дегидрогеназ, с последующим высвобождением энергии в ходе окислительно-восстановительных реакций, где участвуют дыхательные ферменты. Энергия окисления, выделяющаяся в реакциях дегидрогенирования, сохранятся в соединениях водорода с коферментами НАД или ФАД. Для ресинтеза АТФ она используется при переносе водорода от коферментов НАД и ФАД на кислород по системе дыхательных ферментов, которая расположена на внутренней мембране митохондрий.
В дыхательной цепи имеется три пункта сопряжения, где за счет энергии, высвобождающейся при переносе электронов, может синтезироваться АТФ. Первый пункт сопряжения находится на участке переноса водорода от НАД к ФАД. Количества свободной энергии, высвобождаемой в этой реакции, достаточно для синтеза 1 моля АТФ. Второй пункт сопряжения локализуется на участке переноса электронов от кофермента Q через цитохром к цитохрому с. Третья молекула АТФ синтезируется на завершающем этапе дыхательной цепи в цитохромоксидазной реакции, где происходит перенос электронов с системы цитохромов на кислород. В этом пункте сопряжения энергии освобождается больше, чем при любой другой реакции дыхательной цепи. Этой энергии вполне могло бы хватить для образования нескольких молей АТФ, но, тем не менее, в этом пункте сопряжения, как и в первых двух, синтезируется только 1 моль АТФ. Излишек энергии, выделяемой в цитохромоксидазной реакции, служит главной движущей силой для всего процесса переноса электронов по дыхательной цепи. Кислород, акцептирующий электроны, которые поставляет цитохромоксидазная реакция, служит резервуаром, обеспечивающим постоянный отток электронов из дыхательной цепи и поддерживающим ее компоненты в окисленном состоянии, благодаря чему они могут принимать водород от различных субстратов окисления.
Наряду с обычным путем окисления субстратов на внутренней мембране мышечной клетки существует также путь окисления, локализованный на внешней мембране, в котором принимают участие цитохром с, система ФП5 — цитохром Ь5 и цитохромоксидаза. Активация этого пути приводит к быстрому окислению внемитохондриального НАД-Н, но он не связан с синтезом АТФ и ведет к рассеиванию энергии в виде тепла. Этот путь используется в качестве буферной системы, поддерживающей необходимую концентрацию окисленной формы НАД в саркоплазме и устраняющей избыток молочной кислоты, образующейся при гликолизе.
Общая схема, иллюстрирующая взаимосвязь аэробных и анаэробных превращений в энергетическом обмене скелетной мышцы, представлена на рис. 22.1.
Общий выход энергии при аэробном процессе более чем в 10 раз превышает изменение свободной энергии при гликолитическом распаде углеводов в анаэробных условиях. В качестве субстратов аэробных превращений в работающих мышцах могут быть использованы не только внутримышечные запасы гликогена, но и внемышечные резервы углеводов (например, гликоген печени), жиров, а в отдельных случаях и белков. Поэтому суммарная емкость аэробного процесса очень велика и трудно поддается точной оценке. В отличие от гликолиза, метаболическая емкость которого в значительной степени зависит от изменений внутренней среды вследствие накопления в организме избытка молочной кислоты, конечные продукты аэробных превращений — СО2 и Н2О — не вызывают каких-либо
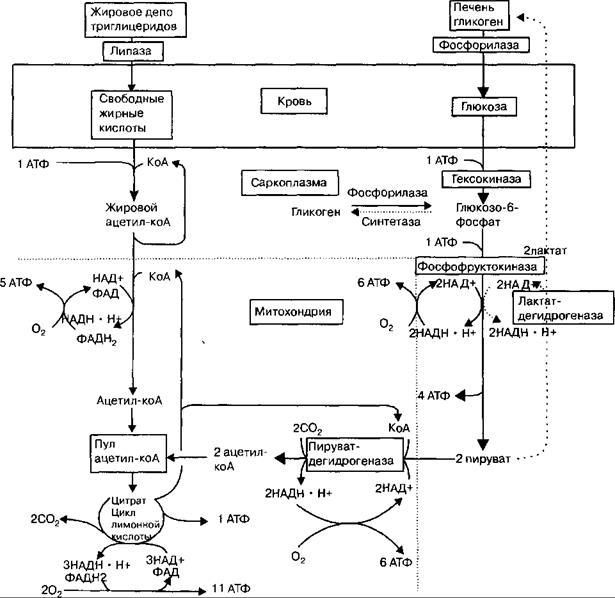
Рис. 22.1. Общая схема образования энергии в мышечных клетках.
Источниками образования АТФ в мышечных клетках являются окислительное фосфорилирование, а также фосфогенная система «гликоген—молочная кислота». Аэробное дыхание является наиболее эффективным, так как из каждой молекулы глюкозы образуется 38 молекул АТФ, а при анаэробном дыхании — только две.
значительных изменений внутренней среды и легко удаляются из организма.
Образование 1 моля АТФ в процессе окислительного фосфорилирования эквивалентно потреблению 3,45 л О2. Столько же кислорода в покое потребляется в течение 10—15 мин, а. при напряженной мышечной деятельности (например, во время бега на марафонскую дистанцию) — за 1 мин. Однако в самих работающих мышцах запасы кислорода крайне невелики. Небольшое его количество находится в растворенном состоянии во внутриклеточной саркоплазме и в связанном состоянии с миоглобином мышц. Основное же количество кислорода, потребляемого в мышцах для ресинтеза АТФ, доставляется в ткани через систему легочного дыхания и кровообращения.
Для бесперебойной работы дыхательной цепи и механизма окислительного фосфорилирования напряжение О2 в клетках должно поддерживаться на уровне не ниже 5—10 мм рт. ст. Чтобы обеспечить его, напряжение О2 снаружи (в мышечных капиллярах) должно быть примерно 15—20 мм рт. ст., поскольку кислород поступает в клетки путем диффузии.
Поддержание критического напряжения О2 на наружной клеточной мембране независимо от изменений скорости расхода кислорода в тканях осуществляет сложная система регуляции, в которую наряду с внутриклеточными механизмами метаболического контроля входят также нервная и гормональная регуляция внешнего дыхания, центрального и периферического кровообращения.
Максимальная мощность аэробного процесса в равной мере зависит как от скорости утилизации О2 в клетках (а она, в свою очередь, от общего числа митохондрий в клетке, количества и активности ферментов аэробного окисления), так и от скорости поставки О2 в ткани.
22.1.3. «Кислородный каскад» и эффективность транспорта кислорода к работающим мышцам
Скорость доставки кислорода к тканям является одним из важнейших факторов, определяющих возможность обеспечения энергией работающих мышц. В митохондриях скелетных мышц, где образуется до 90 % всей необходимой энергии, скорость ресинтеза АТФ находится в зависимости от достигнутой концентрации или напряжения кислорода в клетке. Общая скорость ресинтеза АТФ, обеспечиваемая различными источниками энергии, является функцией напряжения кислорода в клетке. При низком уровне метаболических превращений в клетке, как это имеет место в покоящейся мышце, изменения в скорости доставки О2 в ткани не оказывают влияния на скорость ресинтеза АТФ (зона насыщения). Однако при снижении рО2 в клетке ниже некоторого критического уровня поддержание скорости ресинтеза АТФ на необходимом уровне возможно только за счет адаптивных сдвигов клеточного метаболизма, связанных с природой используемых субстратов, размерами их внутриклеточных запасов, с состоянием фосфорилирования и величиной внутриклеточного pH, что с неизбежностью потребует увеличения скорости доставки О2 в ткани. Максимальная скорость потребления О2 митохондриями скелетных мышц может быть поддержана только до определенного критического значения рО2 в клетке, которое составляет от 0,5 до 3,5 мм рт. ст. Если уровень метаболической активности при мышечной работе превысит значение максимально возможного усиления аэробного ресинтеза АТФ, то возрастающая энергетическая потребность в этих условиях может быть компенсирована за счет анаэробного ресинтеза АТФ. Однако этот диапазон анаэробной метаболической компенсации весьма узок, и выше значения максимального увеличения скорости ресинтеза АТФ, обеспечиваемого сочетанной метаболической активностью аэробного и анаэробных процессов в работающей мышце, дальнейшее исполнение ее сократительной функции становится невозможным. Диапазоны метаболической активности, в пределах которых текущая поставка О2 недостаточна для поддержания необходимого уровня ресинтеза АТФ, обычно обозначаются как гипоксические состояния (т. е. состояния кислородной недостаточности) разной тяжести.
Для того чтобы поддерживать напряжение О2 в митохондриях на уровне, превышающем критическое значение, при котором еще сохраняются условия для адаптивной регуляции клеточного обмена, напряжение О2 на
 |








