2020-06-08
2020-06-08 196
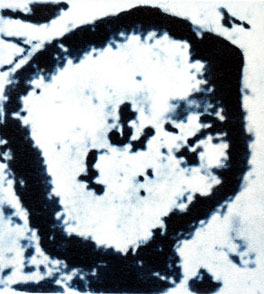
196Что же представляли собой археоциаты как живые организмы далекого геологического прошлого? В 1948 г., на основании следов строения мягких тканей археоциат из кембрия р. Кии в Кузнецком Алатау, удалось доказать, что археоциаты имели не наружный скелет, а внутренний. Губчатое строение центральной полости в целом было принято за внутренний орган ассимиляционного назначения. Эта находка позволила выяснить назначение центральной полости, помогла установить роль пор стенок, перегородок и днищ и, что особенно важно,‑ внутренний скелет у этой группы организмов. Наружная мягкая оболочка прекрасно объясняла, почему археоциаты обладали способностью прикрепляться к субстрату, образуя те или иные дополнительные элементы скелета до отложения иногда сплошной массы твердого скелетного вещества.

Эпифитоны ‑ микроскопические красные водоросли кембрия. Фото автора
Находки археоциат с остаточными структурами мягких тканей внутри кубка и на его поверхности в СССР уже перестали быть редкостью. В южной части Сибири, Восточном Забайкалье среди остатков скелетов археоциат попадаются и такие, у которых в разной степени сохранились следы строения мягких тканей. В некоторых шлифах можно проследить замечательные картины биологических взаимоотношений форм, принадлежащих разным видам и являющихся самостоятельными организмами. Своеобразно выглядело дно кембрийского моря, где, кроме водорослей, появились археоциаты, губки, копошились и плавали разнообразные древнейшие ракообразные ‑ трилобиты.
Самое значительное, что принесло изучение археоциат,‑ это обнаружение их остатков со следами строения мягких тканей. Причина этого, по‑видимому, в отсутствии в водах кембрийских морей гнилостных бактерий. У многих экземпляров археоциат во внутренней полости сохранились следы сложно построенной системы взаимосвязанных каналов, сопряженных с порами внутренней стенки. Сюда поступала морская вода с пищевыми веществами. Продукты ассимиляции перемещались затем по капиллярным сосудам в межстенное пространство, где имелись многочисленные камеры со стенками, покрытыми слоем мягкой ткани. Сквозь поры мягкой наружной оболочки проходили наружу тонкие капиллярные сосуды.
Полученные микрофотографии со следами строения мягкого тела археоциат имеют исключительную ценность для дальнейшего изучения этого интересного типа животных, поздние представители которого к девону совершенно вымерли.
Наряду с археоциатами в кембрии появилась большая группа древнейших членистоногих ‑ трилобиты, эволюция которых растянулась на несколько геологических периодов ‑ до триаса включительно (570‑185 млн. лет назад). Появились древнейшие панцирные рыбы, строматопороидеи, брахиоподы и т. д. С начала кембрийского периода мир животных начал развиваться бурно, сложно, с проявлением многих этапов формообразования, с вымиранием одних групп и появлением новых.
За последнее время в отложениях кембрия обнаружены представители многих новых классов. Автором были установлены криброциаты, сопутствующие почти повсеместно археоциатам, остатки фораминифер в образцах из нижнего кембрия р. Ирбитей на южном склоне хребта Восточного Танну‑Оли, встреченные ранее Е. А. Рейтлингер в синской свите р. Синий, левого притока р. Лены. В образцах своеобразного пятнистого известняка, формировавшегося явно в условиях прибрежно‑морского мелководья, собранных в отложениях ирбитейской свиты на южном склоне хребта Танну‑Ола совместно с археоциатами и губками, и были обнаружены остатки диатомообразных и фораминифероподобных организмов. К. Б. Корде описала новый класс кишечнополостных животных ‑ Hydroconozoa, являющихся породообразующими организмами.
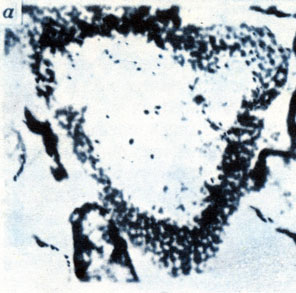
Организмы кембрийского возраста: диатомообразные (ув. в 60 раз). Тува, хр. Танну‑Ола, ирбитейская свита (а); фораминифероподобные (ув. в 210 раз). Тува, хр. Танну‑Ола, р. Кадый, нижний кембрий (б)
Организмы кембрийского возраста: диатомообразные (ув. в 60 раз). Тува, хр. Танну‑Ола, ирбитейская свита (а); фораминифероподобные (ув. в 210 раз). Тува, хр. Танну‑Ола, р. Кадый, нижний кембрий (б)
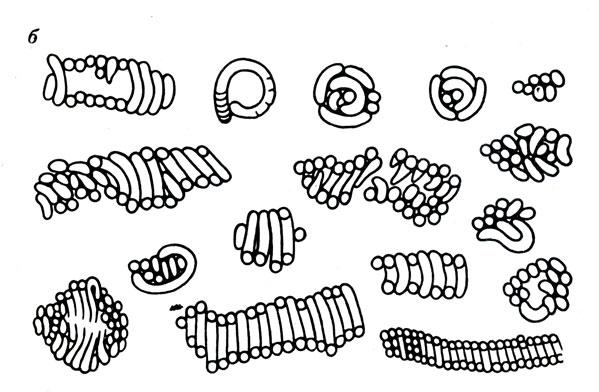
Организмы кембрийского возраста: диатомообразные (ув. в 60 раз). Тува, хр. Танну‑Ола, ирбитейская свита (а); фораминифероподобные (ув. в 210 раз). Тува, хр. Танну‑Ола, р. Кадый, нижний кембрий (б)
Начиная с кембрия стали интенсивно развиваться красные и зеленые (сифоновые) водоросли. Первые были представлены большим количеством родов и видов. Наряду с археоциатами они играли породообразующую роль в биогермах кембрийского возраста, могли сопутствовать археоциатам, но в некоторых случаях были единственными породообразователями,
Остатки красных водорослей встречаются в отложениях морских фаций, сформированных в чистых водах и при нормальной для кембрийского времени солености. Сифоновые водоросли были представлены незначительным количеством видов и родов, но местами ‑ в Казахстане, Сибирской платформе и на р. Амге, они играли породообразующую роль. Синезеленые водоросли с начала кембрия стали уменьшаться в количестве.
Рассматривая комплекс живых существ на Земле как "живое вещество биосферы", следует отметить, что это вещество выполнило в течение палеозоя грандиозную работу. Непрерывно и бурно развиваясь качественно, давая новые и новые формы, оно уже с начала эры стало решительно распространяться и на области суши. Первыми, освоив прибрежные зоны бассейнов, вышли на сушу микроскопические синезеленые водоросли, которые затем довольно быстро распространялись на площадях увлажнения почв. Водоросли с более высокой организацией, красные и зеленые, располагавшие корневой системой и специализацией клеточных тканей, смогли при этом выйти в субаэральную среду. Несомненно, все присущие наземной растительности особенности ‑ анатомическое строение, смена поколений в цикле индивидуального развития и другие ‑ все это было заложено еще в предшествующей, водной, стадии развития.
Выход растений на сушу заставил их приспособиться к самым разнообразным условиям обитания. Поскольку ни свет, ни влага, ни углекислота не лимитировали развития растений, то они быстро расселились на обширных пространствах, одновременно приобретя многие новые специфические черты строения. Ботаники считают, что при этом, кроме ранее господствовавшего полового размножения растений, проявилось и стало развиваться бесполое ‑ вегетативное. Переход на сушу вызывал возникновение у растений способности к защите от потери влаги и высыхания, развитие тканей, которые должны были играть роль опорного каркаса и т. д.

Часть поверхности водорослевого рифа ‑ биогерма. Нижний ордовик. Реконструкция автора
В истории палеозоя были этапы массового развития растений. В условиях влажного климата это приводило местами к накоплению растительного вещества в составе осадочных отложений с последующим превращением его в результате вторичных процессов в каменный уголь. В водных условиях усиливалась жизнедеятельность водорослей, что также приводило к накоплению органических веществ с частичным их преобразованием потом, в толще накапливаемых минеральных осадков, в нефть и горючий газ. По‑видимому, стимулом для подобных усилений жизнедеятельности фотосинтезирующих организмов были выбросы в биосферу, уже значительно обедненную углекислотой, новых ее запасов.