 2020-09-24
2020-09-24 116
116а) компоненты суточных энерготрат организма.
валовый обмен - общая величина энерготрат организма за сутки.
валовый обмен=ОО+РП+СДДП.
б) основной обмен, факторы определяющие величину ОО.
ОО - минимальный уровень энерготрат организма в условии физ. и эмоционального покоя.
состояние ОО: утро, натощак, лежа на спине, состояние спокойного бодрствования, t в помещении 18-22.
Факторы определяющие ОО: генотип, возраст, пол, масса, рост, хар-р питания, состояние н.с. и эндокринной сис-мы.
правило поверхности тела: уровень энерготрат прямопропорционален площади поверхности тела.
Это правило справедливо для вычисления ОО.
в) Специфически-динамическое действие пищи (СДДП).
СДДП - увеличение энерготрат организма после приёма пищи. Белки на 30%, углеводы и жиры по 15%.
г) величины энергетического обмена в пяти осн. профессиональных группах людей.
Рабочий обмен (РО) - величина энерг. затрат для определённого вида трудовой деятельности.
Рабочая прибавка. РП=РО-ОО.
1) умственный труд - 2500-2800
2) полностью механизированный физ. труд - 2800-3500
3) частично механизированный труд - 3500-4000
4) немеханизированный труд - 4000-5000
5) очень тяж. физ. труд - 5000-7000.
Билет9
Физиология вкуса
а) морфофункц. хар-ка периферического, проводникового и коркового отдела вкусового анализатора
Периферический: Вкусовая почка имеет колбовидную форму и не достигает поверхности слизистой оболочки языка и соединена с полостью рта через вкусовую пору.
Проводящие пути и центры вкуса. Проводниками всех видов вкусовой чувствительности служат барабанная струна и языкоглоточный нерв, ядра которых в продолговатом мозге содержат первые нейроны вкусовой системы. Вкусовые афферентные сигналы поступают в ядро одиночного пучка ствола мозга. От ядра одиночного пучка аксоны вторых нейронов восходят в составе медиальной петли до дугообразного ядра таламуса, где расположены третьи нейроны, аксоны которых направляются в корковый центр вкуса.
б) механизм возбуждения вкусовых рецепторов
Вкусовой рецептор, возбуждается благодаря взаимодействию молекул стимулирующего в-ва с рецепторами локализованными в мембране сенсорной вкусовой клетки. В результате происходит конформация рецептора которая приводит к изменению проницаемости клетки и генерации рецепторного потенциала.
в) топография вкусовых рецепторов языка
Вкусовые почки — рецепторы вкуса — расположены на языке, задней стенке глотки, мягком небе, миндалинах и надгортаннике. Больше всего их на кончике, краях и задней части языка.
1. сладкий - кончик языка, 2. солёный - по бокам и спинка, 3. горький - корень.
г) методы исследования вкусовой чувствительности
Поэтому 20 % раствор сахара воспринимается как максимально сладкий,
10 % раствор натрия хлорида — как максимально соленый,
0,2 % раствор соляной кислоты — как максимально кислый, а
0,1 % раствор хинина сульфата — как максимально горький.
Физиология дыхания
а) принципы регуляции дыхания:
- поддержание оптимального парц. напряжения О2 и СО2 в организме за счет управления МОД.
- поддержание оптимального соотношения между частотой и глубиной вентиляции легких.
- приспособление вентиляции лёгких к экзогенным и эндогенным факторам.
б) хар-ка морфофункциональной организации дых. центра.
- корковый отдел
- лимбический
- гипоталамический
- пневмотаксический центр варолиева моста (медиальное парабрахиальное ядро и ядро Шатра)
- гаспинг центр
- дых. центр продолговатого мозга
- спинальные мотонейроны дых. мышц.
Дых. центр продолг. мозга (на дне 4 желудочка в ромб. ямке) состоит из дорсомедиальной (инспир. отдел) и вентролатеральной (ростральная и каудальная (экспир. отдел) части) областей.
В н.у. дых. центр получает афферентные сигналы от периферических и центральных хеморецепторов, сигнализирующих о рО2 в крови и концентрации Н+ во внеклеточной жидкости мозга. Афферентные сигналы от хеморецепторов тесно взаимодействуют с другими афферентными стимулами дых. центра, но, в конечном счете, химический, или гуморальный, контроль дыхания всегда доминирует над нейрогенным.
функции:
- двигательная: генерация дыхательного ритма (вдох и его прекращение); паттерн дыхания - длит-ть вдоха и выдоха, величин ДО, МОД.
- гомеостатическая: поддерживает норм. величины дыхательных газов (O2, CO2) и рН в крови и внеклеточной жидкости мозга, регулирует дыхание при изменении температуры тела, адаптирует дыхательную функцию к условиям измененной газовой среды.
в) класс-я нейронов дых. центра продолговатого мозга:
- ранние инспираторные
- поздние инспираторные
- полные инспираторные
- постинспираторные
- экспираторные
- преинспираторные.
Нейроны дых. центра в зависимости от проекции их аксонов: 1) нейроны, иннервирующие мышцы верхних дых. путей и регулирующие поток воздуха в дых. путях; 2) нейроны, которые синаптически связаны с дых. мотонейронами с.м. и управляют мышцами вдоха и выдоха; 3) проприобульбарные нейроны, которые связаны с другими нейронами дых. центра и участвуют только в генерации дых. ритма (ранние инспираторные и постинспираторные)
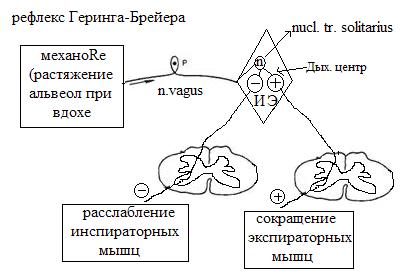
г) блок-схема рефлексов саморегуляции дыхания с механорецепторов альвеол (рефлекс Геринга-Брейера) и с проприорецепторов дых. мышц.????????????
Рефлекс Геринга — Брейера контролирует глубину и частоту дыхания. У человека он имеет физиологическое значение при дыхательных объемах свыше 1 л.
Проприоцептивный контроль дыхания. Межреберные мышцы, в меньшей степени диафрагма, содержат большое количество мышечных веретен. Активность этих рецепторов проявляется при пассивном растяжении мышц, изометрическом сокращении и изолированном сокращении интрафузальных мышечных волокон. Рецепторы посылают сигналы в соответствующие сегменты с.м.. Недостаточное укорочение инспираторных или экспираторных мышц усиливает импульсацию от мышечных веретен, которые через гамма-мотонейроны повышают активность альфа-мотонейронов и дозируют таким образом мышечное усилие.
Билет 10








