 2020-09-24
2020-09-24 205
205Содержание
Введение
1.Общая характеристика транспорта веществ через мембрану
1.1 Пассивный транспорт
1.2 Активный транспорт
2.Каналы
2.1 Никотиновый ацетилхолиновый рецептор
2.2 Натриевый канал
2.3 Кальциевый канал
2.4 Щелевые контакты
3.Поры
3.1 Ядерные поровые комплексы
3.2 Порины
3.3 Мембранные поры, создаваемые экзогенными агентами
4.Молекулы, используемые в качестве моделей пор и каналов
Заключение
Литература
Введение
Живая клетка - это элементарная ячейка биологической организации, обеспечивающая все функции организма. Среди многообразных явлений, протекающих в клетке, важное место занимают активный и пассивный транспорт веществ. В настоящее время стало очевидно, что эти явления, так или иначе, определяются барьерными свойствами клеточных мембран. Клетка - открытая система, которая непрерывно обменивается с окружающей средой веществом и энергией. Во многих случаях биологического транспорта основного переноса веществ является их диффузия через клеточную или многоклеточную мембрану. Способы диффузионного переноса многообразны: диффузия жирорастворимых веществ через липидную часть мембраны, перенос гидрофильных веществ через поры, образуемые мембранными липидами и белками, облегченная диффузия с участием специальных молекул-переносчиков, избирательный транспорт ионов через ионные каналы.
Цель моей работы – изучить строение и свойства проводящих путей биологических мембран.
Для реализации этой цели были поставлены следующие задачи:
1. Ознакомиться с литературой;
2. Изучить особенности пор и каналов;
3. Изучить основные модели биологических структур.
ОБЩАЯ ХАРАКТЕРИСТИКА ТРАНСПОРТА ВЕЩЕСТВ ЧЕРЕЗ МЕМБРАНУ
ПАССИВНЫЙ ТРАНСПОРТ
Под пассивным транспортом веществ через биологические мембраны подразумевается движение молекулы вещества в комплексе с переносчиком. Такой путь проникновения веществ через мембрану не требует затраты энергии, что и дало возможность назвать его пассивным. Для пассивного транспорта характерен эффект насыщения - по мере увеличения концентрации вещества скорость его трансмембранного движения достигает некоторого предела. Так как эффект насыщения очень четко наблюдается также в ферментативных реакциях, в связи с движением в мембране гипотетических переносчиков.
В настоящее время имеется большое количество различных моделей пассивного транспорта. Все они могут быть разделены на две большие группы: модели переносчиков и модели пор. Основное отличие между этими группами моделей заключается в том, что в моделях переносчиков каждый специфический связывающий центр в течение транспортного цикла бывает попеременно доступен омывающим растворам мембраны. В моделях пор каждый специфический центр фиксирован в мембране и доступен или обоим омывающим растворам (одноцентровые поры), или только одному из них, либо вообще недоступен растворам.
Транспорт в порах осуществляется в результате последовательного связывания вещества специфическими центрами. Транспорт же по типу переносчиков осуществляется в результате попеременного экспонирования центров связывания последнего к омывающим мембрану растворам.
АКТИВНЫЙ ТРАНСПОРТ
В последнее время достигнуты большие успехи в изучении активного транспорта, представляющего наибольший интерес из всех видов трансмембранного движения веществ. Особенностью активного транспорта является перенос молекул вещества через мембрану против градиента концентрации, т.е. при неравенстве концентраций (Се>Сi или Сi>Се) существует равенство потоков Iе= Ii. Процессы активного транспорта весьма сложны и протекают с участием нескольких мембранных компонентов.
К активному транспорту веществ через мембраны относятся явления эндо - и экзоцитоза.
Эндоцитоз - это явление, благодаря которому внеклеточные вещества попадают внутрь клетки, заключенные в пузырьки из плазматической мембраны, что часто называют – гетерофагосомами. В этом процессе принимают участие плазматическая мембрана, микрофиламенты и контрактильные нити клетки.
Экзоцитоз - это процесс обратный эндоцитозу. Так удаляются в окружающее клетку пространство остаточные тельца.
КАНАЛЫ
Под каналами чаще всего понимают ионные каналы, которые, как теперь известно, широко распространены во многих типах клеток. Регулируемые ионные каналы, участвующие в передаче сигнала, в ответ на определенный внешний стимул быстро изменяют мембранную проницаемость для определенного иона. При этом происходит изменение трансмембранного потенциала. К работе такого вида каналов предъявляются некоторые требования. Во-первых, внешний сигнал должен вызывать быстрое переключение между открытым и закрытым состояниями канала. Необходимо чтобы быстро устанавливалось равновесие. Когда канал открыт, через бислой может проходить до 106-108 ионов в секунду. Каналы обладают селективностью, т.е. способность канала пропускать некоторые ионы лучше, чем другие; ее можно качественно охарактеризовать как отношение проницаемостей или проводимостей для сравниваемых ионов. Например, у входа в канал имеются отрицательные заряды, то скорость транспорта анионов может уменьшаться, т.е. канал окажется катионселективным. Размеры переносимых веществ также влияют на скорость транспорта.
Существует большое количество видов каналов. Две большие группы составляют потенциалзависимые ионные каналы и каналы, регулируемые нейромедиаторами.
НИКОТИНОВЫЙ АЦЕТИЛХОЛИНОВЫЙ РЕЦЕПТОР (nAChR-канал)
nAChR-канал является примером канала, работа которого регулируется нейромедиаторами. Эти каналы находятся главным образом в концевых пластинках постсинаптических мембран нервно-мышечных соединений. При возбуждении нейрона из него высвобождается нейромедиатор ацетилхолин. При взаимодействии с ацетилхолином канал открывается, опосредуя селективное перемещение катионов. nAChR-канал регулируется с помощью химического механизма, которым является ацетилхолин.
Выделенный и очищенный nAChR-канал состоит из пяти полипептидных субъединиц четырех разных типов со стехиометрией α2βδγ. Две копии α-субъединиц, присутствующие в комплексе, выполняют разные функции. Все субъединицы фосфорилированы и гликозилированы, а к двум α и β, ковалентно присоединен липид. Была построена модель канала, приведенная на рис.2.
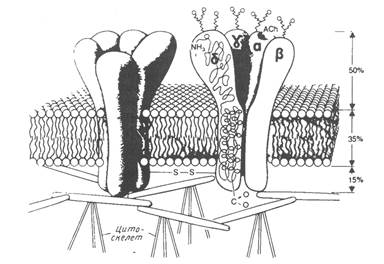
Рис.2. Модель канала никотинового ацетилхолинового рецептора.
Канал имеет центральное отверстие диаметром 30Ǻ с внеклеточного конца и более узкое с цитоплазматической стороны. Пять субъединиц расположены в следующей последовательности: β-α-δ-γ-α, так что α-субъединицы не соседствуют друг с другом. Места связывания располагаются на α-субъединицах. Канал имеет длину около 140Ǻ, причем участок длиной 70Ǻ расположен над поверхностью бислоя с наружной стороны, образуя большие ворота канала. В открытой конформации канал проницаем для катионов и небольших неэлектролитов, но не анионов. Селективность канала по отношению к одно- и двухвалентным катионам невелика. Хотя размеры nAChR-канала относительно велики, он все же слишком мал, чтобы через него могли полностью проходить гидратированные ионы.
НАТРИЕВЫЙ КАНАЛ
Является потенциалзависимым ионным каналом, который обеспечивает быстрое увеличение натриевой проводимости, ответственное за фазу деполяризации при развитии потенциала действия в нервных и мышечных клетках. Каналы, выделенные из тканей млекопитающих, имеют молекулярную массу ~335000. Na+ -каналы взаимодействуют с различными токсинами, в частности с тетродотоксином, сакситоксином и α-токсином скорпиона, которые очень прочно связываются с канальными белками и могут использоваться при количественных биохимических измерениях.
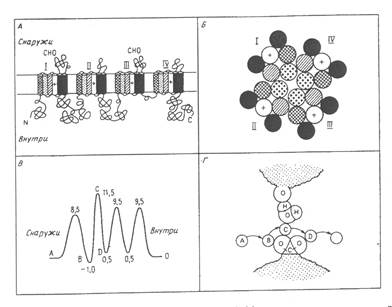
Рис.3. Некоторые модели натриевого канала.
А-модель укладки полипептидной цепи; показаны четыре гомологичных домена, каждый из которых имеет шесть трансмембранных спиралей. Знаком + отмечен амфифильный сегмент.Б-гипотетическая организация шести спиралей каждого домена, образующих центр.канал.В-профиль свободной энергии.Г-гипотетическая модель узкого «селективного фильтра».
Было показано, что белок натриевого канала содержит 1820аминокислотных остатков, организованных в четыре повторяющиеся единицы (рис.3.). Каждый гомологичный участок содержит 4, 6 или 8 трансмембранных α-спиралей. Некоторые из этих трансмембранных спиралей амфифильны. Специфические группы организованы в канале таким образом, что образуется «селективный фильтр».
На рис.3. приведен профиль свободной энергии для диффузии ионов Na+ через канал. Если лимитирующей стадией является дегидратация, то параметром, определяющим высоту энергетического барьера, будет геометрия тех компонентов, которые временно заменяют воду (пик В на рис.3.).
КАЛЬЦИЕВЫЙ КАНАЛ
Ca2+-селективные каналы так же относятся к потенциалзависимым ионным каналам. Широко распространены в возбудимых клетках - нервных и мышечных, а также в большинстве других типов клеток. Некоторые Ca2+ -каналы отвечают на изменение напряжения на мембране. Обычно концентрация ионов Ca2+ в цитоплазме не превышает 10М, что в 10000 раз ниже, чем концентрация ионов Ca2+ вне клетки. Открывание Ca2+--канала может приводить к значительным изменения концентрации этого иона в цитоплазме, что в свою очередь индуцирует разнообразные биохимические события. Каналы состоят как минимум из двух субъединиц с молекулярной массой 140000 и 30000. Большая субъединица была клонирована и секвенирована, и оказалось структурно близка к потенциалзависимому натриевому каналу.
ЩЕЛЕВЫЕ КОНТАКТЫ
Щелевые контакты - это кластеры мембранных каналов, которые соединяют содержимое соседних клеток в тканях. Через такие каналы проходят небольшие молекулы - метаболиты и неорганические ионы. Диаметр каналов в клетках млекопитающих составляет от 12 до 20Ǻ. Эти каналы соединяют две плазматические мембраны. Исходя из данных об аминокислотной последовательности, можно предположить, что в каждой субъединице имеется четыре трансмембранные α-спирали. Эти каналы находятся обычно в открытом состоянии, но закрываются, когда понижается скорость метаболизма. Каждый канал состоит из 12 субъединиц, по шесть от каждой клетки. Канал представляет собой гексамерную структуру. Два гексамерных комплекса соседних мембран соединены конец к концу и образуют протяженный канал, объединяющий обе мембраны. Структура канала щелевого контакта зависит от наличия ионов Ca2+ . В присутствии Ca2+ субъединицы расположены параллельно центральной оси канала, а в отсутствии этих ионов несколько наклонены (открытое состояние). Точный механизм открывания и закрывания далеко не ясен.
ПОРЫ
В последнее время достигнуты большие успехи в определении строения пор на молекулярном уровне. Особенно ценным в исследованиях оказался метод реконструкции изображения; с его помощью удалось не только визуализировать отверстия в мембране, создаваемые большими порами, но и выявить симметричную организацию субъединиц вокруг центрального отверстия (табл.2).
Таблица 2. Псевдосимметрия некоторых пор.
| Пора | Структурная симметрия | Комментарии |
| Порины | Симметрия 3-го порядка | Три идентичные субъединицы |
| Ядерный поровый комплекс | Симметрия 8-го порядка | Субъединичный состав неизвестен |
Важным исключением из α-спирального семейства являются порины, поскольку они формируют поры из β-слоев, а не с помощью α-спиралей. Поры могут образовываться с помощью эндо- и экзогенных веществ.








