 2020-09-24
2020-09-24 388
388
Ядерная оболочка клеток млекопитающих содержит 3-4 тысячи пор (примерно 10 пор на 1 квадратный мкм). Через ядерные поры происходит обмен веществами между ядром и цитоплазмой. Действительно, РНК, синтезируемые в ядре, а также рибосомные субъединицы и белки, содержащие сигналы ядерного экспорта, транспортируются через ядерные поры в цитоплазму, а гистоны, компоненты репликативной системы, многие другие белки импортируются через ядерные поры из цитоплазмы в ядро. Поры окружены большими кольцевыми структурами, называемыми поровыми комплексами (их внутренний диаметр составляет приблизительно 80 нм, а мол. масса -50-100 млн. Каждый комплекс образован набором больших белковых гранул, сгруппированных в октагональную структуру. Поровой комплекс пронизывает двойную мембрану, связывая по окружности поры липидный бислой внутренней и внешней мембран в единое целое. "Дыра" в центре каждого комплекса (ядерная пора) представляет собой водный канал, сквозь который водорастворимые молекулы курсируют между ядром и цитоплазмой. Ядерный поровой комплекс содержит заполненный водой цилиндрический канал диаметром около 9 нм. Большие ядерные белки взаимодействуют с белками-рецепторами, расположенными на границе ядерных пор, и эти рецепторы активно переносят белки в ядро, увеличивая канал поры.
Количество ядерных пор зависит от типа клетки, стадии клеточного цикла и конкретной гормональной ситуации. Для ядерной поры характерна симметрия восьмого порядка, поэтому многие белки ядерной поры представлены в ее составе в количестве, кратном восьми. В электронный микроскоп видны выпуклые кольца. Кольцо, находящееся с ядерной стороны, несет структуру, называемую корзиной (basket). Это образование состоит из обращенных в нуклеоплазму фибрилл и прикрепленного к ним терминального кольца. К просвету канала обращены восемь симметричных образований (spoke complex). В центре комплекса виден вход в канал ядерной поры. Иногда в канале оказывается видна электронноплотная гранула. Некоторые исследователи полагают, что это какой-то транспортирующийся комплекс в момент пересечения ядерной мембраны. Другие считают, что эта структура является функциональной деталью ядерной поры. На основании этого последнего предположения была даже выдвинута не подтвердившаяся впоследствии гипотеза, согласно которой ядерная пора содержит не один, а восемь проницаемых каналов. Молекулы массой менее 5 кДа, проходят через ядерную пору свободно, и равновесие между ядерной и цитоплазматической концентрацией устанавливается за секунды. Для белков массой 17 кДа этот процесс занимает 2 минуты, белков массой 44 кДа (приблизительно 6 нм) - 30 минут. Белки массой более 60 кДа, по-видимому, вообще не могут пассивно проходить через ядерные поры. Проницаемый для гидрофильных макромолекул канал, через который происходит как активный, так и пассивный транспорт, в ядерной поре один, и он, по всей видимости, расположен в центре комплекса. Существуют специальные механизмы транспорта макромолекул внутрь ядра и из ядра в цитоплазму, однако до сих пор о них мало что известно.
ПОРИНЫ
Порины образуют поры, которые функционируют как молекулярные сита, опосредуя диффузию небольших гидрофильных молекул через наружную мембрану грамотрицательных бактерий. Молекулярная масса поринов варьирует от 28000 до 48000. В мембране обычно присутствуют в виде триммеров. Для поринов характерно высокое (до 60%) содержание β-слоев. Наиболее полно к настоящему времени охарактеризованы порины из Escherichia coli: ОmpF (порин матрикса), ОmpС, РhoE, LamB (мальтопорин). Эти белки имеют молекулярную массу ~35000. Их основной особенностью является то, что они образуют наполненный водой трансмембранный канал, причем этот канал образован в основном β-структурами. На рис.4. представлена одна из возможных моделей образования поринового канала из амфифильных β-цепей.
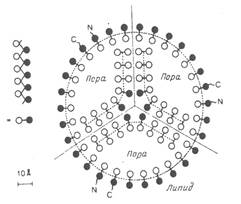
Рис.4.Модель поринового тримера (вид сверху).
Образуемые порином каналы различаются как по размерам (диаметр от 6 до 23Ǻ), так и по селективности. Селективность связана с наличием внутри или около входа заряженных аминокислотных остатков. В одних случаях порины образуют один большой канал, в других – три независимых (рис.4.).
Три из четырех поринов Escherichia coli имеют много общих структурных особенностей, а их аминокислотные последовательности в значительной степени гомологичны. Эти порины образуют поры диаметром 10-12 Ǻ (ОmpF, ОmpС, РhoE).
Порины представляют большой интерес, поскольку они показывают, что трансмембранные каналы могут образовывать не только из α-спиралей, но и из β-слоев.








