 2020-09-27
2020-09-27 217
217Опыт гомеопатии уникален, прежде всего, тем, что в гомеопатической терапии обычно используют сверхмалые дозы. В отличие от микродоз, имеющих концентрацию порядка 10-3-10-6 массовых долей, разведения гомеопатических препаратов обычно находятся в диапазоне от 10-6 до 10-2000 и выше. Несмотря на то, что в соответствии с законом Авогадро, в разведениях свыше 10-24 М отсутствует даже единая молекула исходного вещества, специфичность терапевтического эффекта гомеопатических препаратов и в «молекулосодержащих» концентрациях, и в умозрительно малых концентрациях свыше 10-24 сохраняется.
Другая особенность гомеопатии - максимальная индивидуализация лечения - основана на использовании клинико-феноменологического принципа подобия.
Основы гомеопатии заложены в 1796 г. Самюэлем Ганеманом, объединившим воедино принцип подобия и принцип использования малых доз. К сожалению, в дальнейшем доминирование принципа подобия задержало исследование физиологических свойств малых доз.
И только теперь, обладая определенным багажом экспериментальных результатов, мы понимаем, что С. Ганеман намного опередил свое время, гениально предложив частный случай применения малых доз.
Для образного восприятия основ гомеопатии целесообразно представить схему, по которой Ганеман испытывал гомеопатические препараты. Общеизвестно, например, что токсическое действие настойки мухомора клинически выражается диареей, специфическим дерматитом, миоклониями, зрительными галлюцинациями и эйфорией. Если токсическую дозу настойки мухомора последовательно уменьшать и назначать группе из 100 здоровых (!) испытуемых, то с уменьшением дозы уменьшится и число испытуемых, реагирующих на нее. На каком-то этапе количество респондентов сократится до 2-3 из 100. Удивительно то, что эти люди похожи друг на друга внешне, чертами характера, поведенческими привычками, пищевыми пристрастиями, а также предрасположенностью к тем или иным заболеваниям, в том числе к наследственным. Гомеопаты относят таких людей к единому гомеопатическому типу или конституции - в данном случае, к конституции мухомора (агарикуса). Не менее удивительно и то, что респонденты с конституцией агарикуса однотипно реагируют и на так называемый потенцированный мухомор. В своих опытах Ганеман не остановился на использовании малых доз, а продолжал последовательно разводить исходные настойки в 100, 1000, 10000 раз и т.д. При этом каждое разведение Ганеман сопровождал 10-кратным встряхиванием раствора в течение 10 секунд. Количество разведений он называл потенциями, а полученные лекарства - потенцированными. Например, 50-я сотенная потенция (СН50)* означает, что 10% настойку мухомора (или другого препарата) 50 раз подвергли разведению, и каждый раз 1 каплю вещества разводили в 99 каплях растворителя. Следовательно, условная концентрация вещества в таком гомеопатическом препарате имеет порядок 100-50 массовых долей**. Как видно, понятие дозы в гомеопатии имеет условный характер и означает степень разведения используемого раствора. Дозы, не содержащие молекул, мы условно называем «нематериальными» (равно как и немолекулярные процессы).
* Появление или усиление у пациента токсических симптомов, характерных для какого-либо вещества, при воздействии потенцированного средства, приготовленного из этого вещества, со времен С. Ганемана называют гомеопатическим обострением.
** Сотенное (С) разведение по Ганеману (Н).
Для назначения потенцированного вещества по принципу подобия необходимо совпадение клинической картины болезни конкретного больного и клинической картины отравления здорового человека этим же средством в токсической дозе. Для достижения терапевтического эффекта врач- гомеопат должен также учитывать наличие у пациента индивидуальной чувствительности, т.е. его конституцию. К примеру, если малая доза настойки мухомора вызывает у здорового человека галлюцинации, то потенцированный мухомор устраняет схожие галлюцинации также только у пациента, имеющего индивидуально высокую чувствительность к мухомору:
| Здоровый человек (чувствительный к мухомору) | + мухомор - (малая доза) | галлюцинации |
| Больной человек, страдающий галлюцинациями (чувствительный, по мнению врача, к мухомору) | гомеопатическое + лекарство = агарикус (потенцированный мухомор) | выздоровление |
В гомеопатии традиционно считалось, что без соблюдения принципа подобия потенцированные препараты не могут оказывать не только терапевтического эффекта, но и другого сколько-нибудь значительного действия.
Однако в исследованиях, результаты которых будут изложены далее, было установлено, что любое потенцированное средство, даже в «нематериальных» дозах обладает присущей только ему специфической активностью. Более того, мы показали, что физиологические эффекты вещества в привычной дозе и приготовленного из него потенцированного препарата в качественном плане идентичны. Эти факты позволили нам условно назвать потенцированное вещество информационной детерминантой и с иных позиций рассматривать процесс потенцирования.
При кажущейся простоте приготовления гомеопатических лекарств, потенцирование - последовательное разведение и встряхивание раствора - придает потенцированным препаратам особые свойства. В литературе имеются работы, показывающие, что в одной и той же малой концентрации до 10-24 растворы, подвергшиеся процессу потенцирования, обладают более высокой биологической активностью, чем просто разбавленные растворы (Harisch G., Dittmann J., 1998).
При высоких разведениях, в отличие от потенцированных растворов, у просто разбавленных растворов биологическая активность отсутствует (Benveniste J. et al., 1992).
Механизм процесса потенцирования неизвестен. Наиболее распространенное объяснение потенцирования зиждется на том, что молекулы растворителя ориентируются вокруг растворяемого вещества в определенном (кластерном) порядке. Кластеры своей пространственной конфигурацией фиксируют физические свойства вещества и при дальнейшем разведении раствора сохраняют эту информацию.
Насколько справедлива кластерная гипотеза, покажет время, но в любом случае мы можем предполагать, что потенцированный раствор является структурированным. По сути, процесс потенцирования является «дематериализацией» вещества, переводом его в информационную форму. Именно этот факт позволил нам предположить, что возможен и обратный процесс - перевод «информации» в «вещество», т.е. постулированная нами выше возможность гипостазирования («материализации») информации в трехмерно-пространственные образования. Обязательная необходимость многократного встряхивания (колебания) раствора для сохранения биологической активности указывает на то, что гипостазирование - волновой процесс, что именно колебания вакуума приводят к его «материализации», а сам первичный информационный алгоритм представляет собой некий код колебаний. Следовательно, и образуемые первичным информационным алгоритмом информационные формы обладают конгруэнтными ему пространственно-колебательными характеристиками.
Подобие клинических проявлений видовой чувствительности (однотипная картина интоксикации всех ста испытуемых настойкой мухомора в приведенном выше примере) и индивидуальной чувствительности, коррелирующей с понятием конституция, позволили предположить, что за этим обыденным фактором «скрыты» различные регуляторные системы - горизонтальная и вертикальная регуляция.
Мысль о том, что информационные детерминанты (формы) представляют собой иерархию пространственно-временных взаимоотношений первичных физических факторов, также обусловлена рядом известных в гомеопатии фактов.
Известно, что у одного и того же человека избирательная индивидуальная чувствительность может проявляться на несколько веществ. Причем, выраженность ее проявлений различная, т.е. существует иерархия индивидуальной чувствительности. Кроме того, в гомеопатии известен ряд препаратов, принадлежащих по происхождению к различным классам веществ, но действующих примерно одинаково. Например, минерал сульфат калия и растение пульсатилла (из семейства лютиковых), известная всем белладонна и карбонат кальция. Опытные гомеопаты сравнительно часто назначают «минералы», так как приготовленные из них потенцированные препараты обладают более широким терапевтическим диапазоном, и действуют «глубже», способны позитивно влиять на течение хронических болезней. Растительные же препараты более «эргичны» - лучше помогают при острых состояниях.
Эффекты потенцированных веществ обладают пространственной ориентацией. Например, ряд гомеопатических лекарств имеет «тропность» к одной из сторон тела: лахезис, яд змеи бушмейстер, эффективен при левосторонних гемиплегиях; а ботропс, яд гадюки - при правосторонних.
Примером временной градуированности эффектов потенцированных средств может служить известная всем гомеопатам закономерность, установленная Константином Герингом: излечение происходит в порядке, обратном появлению симптомов, т.е. при назначении гомеопатического лекарства сначала исчезают недавно появившиеся симптомы, а застарелые - в последнюю очередь.
Эффект бипатии
Одним из вариантов гомеопатии является изопатический метод. Состоит он в том, что уже наличествующие симптомы острой или хронической интоксикации каким-либо веществом устраняют назначением этого же вещества в потенцированной форме. В Англии кожные ожоги, экспериментально вызываемые ипритом, например, успешно лечили ипритом в сверхмалых (гомеопатических) дозах, а потенцированный преднизолон традиционно назначается при таком осложнении терапии кортикостероидами, как синдром Иценко-Кушинга.
Однако способ сочетанного введения в организм одного и того же исходного вещества в значимой весовой дозе и в потенцированной форме в доступной литературе не упоминается, впервые был предложен нами и назван бипатическим. Принципиально «бипатия» от изопатии отличается тем, что в последнем случае потенцированная форма вещества, вызвавшего интоксикацию, назначается после прекращения воздействия исходного вещества на организм.
Первый бипатический эксперимент был проведен в лаборатории биофизики Киевского НИИ оториноларингологии под руководством А. Ф. Карася и К. Н. Игруновой, при участии А. И. Розкладки, О. Ф. Мельникова, С. В. Тимченко и др. Методологически он заключался в том, что животным вводили сочетанно (одновременно) преднизолон в значимой весовой дозе и в потенцированной форме СН30, т.е. условная концентрация преднизолона в последнем случае равнялась 10-60 массовых долей.
Первая часть исследования была проведена на 30 крысах: изучалось бипатическое введение преднизолона на модели острого экспериментального воспаления, которое индуцировали путем введения в заднюю лапку животных 0.05 мг 1% раствора карагинина. В одной группе животным с искусственно вызванным воспалением через зонд per os вводили потенцированный преднизолон, в другой - преднизолон в суммарной дозе 20 мг на крысу. В третьей группе оба препарата вводились сочетанно.
Во всех случаях указанные средства вводились дважды - за один час до инъекции карагинина и через два часа после нее. Контролем служили как интактные животные, так и крысы с «чистым» карагининовым воспалением.
Исследования показали, что введение только преднизолона или преднизолона совместно с его потенцированной формой достоверно угнетает развитие воспалительного отека в лапке крыс. Сама по себе потенцированная форма противовоспалительным действием не обладала и не усиливала его.
Однако бипатическое введение сопровождалось рядом позитивных явлений. При этом при воспалении снижалась способность перитонеальных макрофагов к миграции. Просто преднизолон не обладал этим свойством. При сочетанном же введении способность макрофагов к миграции быстро восстанавливалась до контрольных значений.
Электронно-микроскопические и морфологические исследования показали, что после введения 20 мг преднизолона у животных отмечались умеренно выраженные деструктивные изменения в печени и вилочковой железе. Совместное введение этих же доз препарата и потенцированной формы структуру этих органов не нарушало.
Карагининовое воспаление сопровождается повышенными энергетическими затратами организма на «поддержание» защитного воспалительного процесса. Преднизолон сам по себе не влиял на содержание АТФ в крови, но при бипатическом введении уровень АТФ резко снижался, что ограничивало энергообеспеченность воспалительного процесса. Сочетанное введение преднизолона и его сверхмалой дозы также позитивно сказалось на активности ферментов в клетках крови и привело к увеличению в нейтрофилах щелочной фосфатазы, нормализации уровня АТФазы и 5’-нуклеотидазы, восстановлению активности лактатдегидрогеназы, что в целом свидетельствовало о нормализации клеточного энергообеспечения.
По сравнению с изолированным введением преднизолона, при бипатическом способе наблюдалось более выраженная стимуляция биосинтетической активности лимфоцитов периферической крови крыс, о чем можно было судить по повышению количества в них РНК.
Во второй части исследования изучалась возможность использования потенцированной формы преднизолона в том же гомеопатическом разведении - КГ60 масс, долей для предотвращения побочных эффектов значимых доз преднизолона при хроническом (двухнедельном) введении последнего. Преднизолон вводили один раз в сутки зондом per os из расчета 50 мг на 1 кг массы тела (на модели описанного выше каргининового воспаления эта доза оказывала выраженное противовоспалительное действие). Оказалось, что потенцированный препарат оказывал широкий ряд протекторных эффектов, наиболее существенные из которых приведены в табл. 1.
Представленные в таблице 1 результаты свидетельствуют о том, что потенцированный препарат предотвращает нарушение клеточного метаболизма в лимфоцитах и нейтрофилах и, вероятно, оказывает нормализующее влияние на изменения в мембранных процессах, возникающих при воздействии значимых весовых доз преднизолона.
Кроме того, исследования показали, что потенцированная форма преднизолона предотвращает характерные для длительного применения преднизолона деструктивные изменения в печени, надпочечниках, лимфатических узлах, а также эрозию слизистой желудка. Потенцированная форма преднизолона восстанавливает угнетенную длительным приемом преднизолона синтетическую активность лимфоцитов (увеличивает количество ДНК, не связанной с белками хроматина), т.е. стимулирует функциональную активность лимфоцитов.
Таблица 1.
Протекторные эффекты потенцированного преднизолона.
| Преднизолон | Преднизолон + потенцированная форма |
| Значимое повышение количества палочкоядерных гранулоцитов | Незначительное увеличение количества палочкоядерных гранулоцитов. Достоверное снижение абсолютного содержания моноцитов |
| Снижение содержания лейкоцитов по I сравнению с контролем | Содержание лейкоцитов достоверно не отличается от контроля |
| Не влияет на показатели перекисной хемилюминесценции | Активация процессов перекисного окисления в плазме крови |
| Двукратное снижение уровня АТФ в крови | Увеличение содержания АТФ в крови до уровня, превышающего норму |
| Двукратное снижение содержания щелочной фосфатазы в нейтрофильных лейкоцитах | Незначительное снижение уровня щелочной фосфатазы в нейтрофилах |
| Повышение активности лактатдегидрогеназы в нейтрофилах | Активность лактатдегидрогеназы в нейтрофилах практически не изменена |
| Снижение активности АТФазы в лимфоцитах почти в 2 раза | Активность АТФазы в лимфоцитах не изменена |
Полученные данные позволили предположить, что многочисленные позитивные эффекты потенцированного преднизолона по отношению к токсическим дозам преднизолона образуют новый, ранее неизвестный феномен, названный нами феноменом системной адаптации. Поскольку феномен системной адаптации явился следствием сочетанного применения одного и того же вещества в двух формах - потенцированной и молекулярной, то естественно возникла мысль о возможности его универсального характера.
Для проверки данного предположения в Украинском НИИ клинической и экспериментальной неврологии и психиатрии в лаборатории Т. М. Воробьевой было проведено исследование сочетанного применения этанола с его же потенцированными формами СН30 и СН200 (т.е. имеющими условную концентрацию соответственно 10-60 и 10-200 массовых долей). Изучали следующие показатели: электрическая активность мозга, условно-рефлекторная деятельность, характеристики самостимуляции латерального гипоталамуса, структурная организация сна, содержание эндогенных моноаминов, содержание этанола в крови, активность алкоголь дегидрогеназы в печени, морфо-гистологическая структура печени и мозга.
Исследования проведены на 122 белых крысах - по 8-10 животных в каждой серии. Целью исследования было выяснение возможности использования потенцированных форм этанола в качестве противоалкогольных лекарственных средств. Нейробиологический анализ проводился в различных экспериментальных режимах - в условиях острой и хронической алкогольной интоксикации.
Не вдаваясь в подробности исследования, укажем только наиболее демонстративные его результаты, подтверждающие универсальность феномена системной адаптации:
- Через 30 минут после однократного перорального введения крысам 1 мл 45% этанола в гипоталамусе животных проявлялись пароксизмы альфа- и тета-волн высокой амплитуды (до 300 мкВ). При сочетанном введении этанола с его потенцированной формой (как СН30, так и СН200) указанные пароксизмы не вызывались, наблюдалась медленная синхронизированная активность, свойственная состоянию «спокойного» бодрствования.
- При обучении животных на фоне хронической алкоголизации потенцированные формы этанола сохраняли условно-рефлекторную деятельность крыс на уровне, близком к норме. Так, у интактных крыс количество условно-рефлекторных ответов (в % к числу предъявленных условных сигналов) - 57% (норма), а при длительном ежедневном приеме 1 мл 45% этанола снижалось до 24.5%. При сочетанном введении этого же количества алкоголя и потенцированного этанола в дозе СГ30 - 49%, а СН200 - 41%.
- Потенцированные препараты при сочетанном применении со значимой (I мл) дозой этанола оказывали нормализующее влияние на эмоциональное состояние животных (функциональное состояние так называемой мозговой системы эмоционального подкрепления), о чем свидетельствует следующий эксперимент.
Таблица 2.
Динамика частоты реакции самостимуляции вентролатерального гипоталамуса у крыс за 1 мин: в норме, под влиянием этанола, после одновременного воздействия этанола и его потенцированных форм.
|
| Алкоголь | Алкоголь + потенцированный этанол СН30 | Алкоголь + потенцированный этанол СН200 | ||||
| Первые 0,5 ч | Вторые 0,5 ч | 0,5 ч до введения | 0,5 ч после введения | 0,5 ч до введения | 0,5 ч после введения | 0,5 ч до введения | 0,5 ч после введения |
| 43,6±1,5 | 41,1 ±3,7 | 47,2±2,1 | 58,4*±3,8 | 43,0±1,2 | 32,7*±0,8 | ||
| 64,3±4,4 | 39,2±2,3 | 74,8±3,8 | 44,1 *±1,9 | 76,2±3,3 | 44,6*±2,7 | ||
· - при р < 0,05 в сопоставлении с исходными данными.
Животные имели возможность нажатием педали через введенный стереотаксическим способом электрод раздражать «центр удовольствия» в латеральном гипоталамусе. Алкоголь увеличивал число нажатий на педаль при исходно низком уровне и уменьшал при исходно высоком. Потенцированные препараты, как видно из табл. 2, также оказывали «нормогенное» действие на частоту реакций самостимуляции.
- При длительном ежедневном введении 1 мл 45% алкоголя у крыс сокращалось время сна, а в структуре сна уменьшалась доля парадоксальной фазы сна и фазы глубокого медленного сна. При бипатическом введении этанола (Рис. 12) указанные негативные влияния алкоголя нивелируются. При этом резко увеличивается представленность парадоксальной фазы сна, что свидетельствует о мощном адаптивном эффекте потенцированных средств.
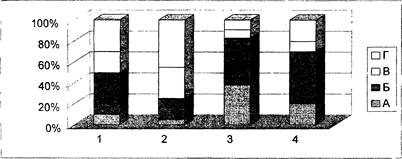
· Рис. 12. Влияние потенцированных форм этанола на структуру цикла бодрствование-сон. 1 - интактные; 2 - после хронической алкоголизации; 3 - после приема потенцированного этанола СН30; 4 - после приема потенцированного этанола СН200. А - парадоксальный сон; Б - глубокий медленный сон; В - поверхностный сон; Г - бодрствование.
- Сложнее ожидаемого оказалось влияние потенцированных препаратов на концентрацию алкоголя в крови: при остром введении его содержание достоверно не изменялось, а при хроническом - увеличивалось (!). При этом наблюдалось повышение активности алкоголь дегидрогеназы.
- На фоне алкоголизации препараты потенцированного этанола изменяли содержание биогенных моноаминов как в мозге, так и в крови (Табл. 3 и 4). Наиболее ярким из адаптивных механизмов при этом является значительное снижение в крови уровня медиатора стресса - адреналина.
- Бипатическое введение алкоголя уменьшало его деструктивное влияние на печень и мозг как при острой, так и при хронической интоксикации.
Таблица 3.
Влияние потенцированного этанола в разведении СН30 и СН200 на содержание биогенных моноаминов (мкг/г ткани) в мозге (гипоталамус + септум) алкоголизированных крыс (М±m; n=10).
| Воздействие | Дофамин | Норадреналин | Адреналин | Серотонин |
| Интактные | 1.10 ± 0.110 | 0.57 ± 0.035 | 0.24 ± 0.023 | 1.64 ±0.072 |
| Однократное введение этанола | 0.66* ±0.109 | 0.53 ± 0.085 | 0.07* ±0.014 | 1.50 ±0.031 |
| Этанол + разведение СH30 | 0.95 ±0.142 | 0.92* ± 0.157 | 0.09* ± 0.021 | 1.97 ±0.258 |
| Этанол + разведение СН200 | 1.08 ±0.062 | 0.98* ±0.137 | 0.10* ±0.012 | 2,17** ±0.092 |
| Хроническое введение этанола | 0.59* ± 0.085 | 0.32* ±0.057 | 0.13* ±0.018 | 1.2* ±0.09 |
| Хроническое введение этанола + разведение СH30 | 0.46* ± 0.099 | 0.14'* ±0.02 | 0.06"* ± 0.003 | 0.71'* ±0.063 |
| Хроническое введение этанола + разведение СН200 | 0.70* ±0.098 | 0.27*°±0.049 | 0.13*°±0.02 | 1 68*°±0.137 |
*- р < 0.05 при сопоставлении с группой «интактные»; # - р < 0.05 при сопоставлении с соответствующей группой алкоголизированных крыс;
**- р < 0.05 при сопоставлении групп крыс, получавших препараты потенцированного этанола в разведении СН30 и СН200.
Таблица 4.
Влияние потенцированного этанола в разведении СН30 и СН200 на содержание биогенных моноаминов (мкг/г ткани) в цельной крови алкоголизированных крыс (М±m; n=10).
| Воздействие | Дофамин | Норадреналин | Адреналин | Серотонин |
| Интактные | 8.20 ±0.62 | 7.74 ±0.69 | 6.00 ± 0.70 | 188.2 ±2.78 |
| Хроническое введение этанола (14 дней) | 12.97* ±2.05 | 12.72*± 2.41 | 16.40* ±2.34 | 151.0 ±28.90 |
| Хроническое введение этанола + разведение СH30 | 8.12* ±1.02 | 5.1* ± 1.11 | 4.79* ± 1.19 | 170.6 ±20.11 |
| Хроническое введение этанола + I разведение СН200 | 10.52 ±1.98 | 6.51*± 1.24 | 5.32* ± 0.98 | 225.9 ± 39.0 |
*- р < 0.05 при сопоставлении с группой «интактные»; # - р < 0.05 при сопоставлении с группой алкоголизированных крыс.
Аналогичные адаптивные механизмы были выявлены при изучении в лаборатории Т. М. Воробьевой бипатического введения животным морфина.
«Бипатические» адаптивные реакции носят сложный характер, о чем свидетельствуют результаты эксперимента, проведенного в НИИ фармакологии МЗ РФ под руководством члена-корреспондента РАМН Б. И. Любимова.
Подопытным животным (мышам) в опытной группе вводили одновременно с токсическими дозами морфина потенцированный морфин СН200 (10-400 масс. долей). В контрольной группе животные получали эти же дозы одного морфина. Итоги эксперимента приведены в табл. 5.
Таблица 5.
Токсичность морфина при его изолированном и «бипатическом» (в комбинации с потенцированным морфином) введении.
| Самки | Самцы | |||
| Доза мг/кг | Количество животных в группе | Количество погибших особей | Количество животных в группе | Количество погибших особей I |
| 150 | 72/72 | 0/7 | 72/70 | 0/7 |
| 175 | 72/72 | 10/24 | 72/70 | 10/29 |
| 200 | 72/72 | 12/24 | 72/70 | 12/26 |
| 250 | 72/72 | 36/32 | 72/70 | 36/30 |
| 300 | 72/72 | 48/42 | 72/70 | 48/35 |
В числителе - количество животных, получивших морфин изолированно (контроль), а в знаменателе - бипатически (сочетание).
Установлено, что:
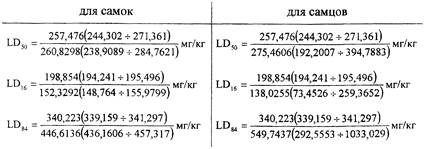
В числителе указан обычный (изолированный), а в знаменателе - бипатический способы введения морфина.
Следующий эксперимент проведен под эгидой фирмы «Материа Медика» в Казанском государственном университете под руководством зав. НИЛ КС д.х.н. профессора А. В. Захарова и старшего научного сотрудника к.х.н. В. Г. Штурлина.
Цель исследования заключалась в количественном изучении влияния процесса потенцирования на кинетику фундаментальной реакции гидролиза АТФ в физиологическом диапазоне pH с цитратным буфером методом ядерно-магнитного резонанса. В одном случае изучалась скорость реакции гидролиза при внесении потенцированной АТФ, в другом - потенцированного буфера. Результаты исследований приведены в табл. 6:
Таблица 6.
Влияние потенцированных форм на реакцию гидролиза.

К - константа скорости гидролиза АТФ в водных растворах эталонных и потенцированных образцов с цитратным буфером при PH 6,7 и Т=378 К;
№№ 1 и 2 - реакции с двумя различными эталонами;
№ 3 - внесено 10% об. потенцированного по правилам гомеопатии АТФ в разведении СН30;
№ 4 - внесено 10% об. потенцированного по правилам гомеопатии нитратного буфера в разведении СН30.
Таким образом, было зафиксировано замедление скорости реакции гидролиза, если один из компонентов использовался в бипатической форме.
Полученные результаты показывают, что бипатический феномен имеет место также в более простых, неорганических системах.
На сегодняшний день наиболее формализованно и достоверно демонстрирует наличие бипатического феномена исследование влияния на синаптическую передачу сверхмалых доз антисыворотки к мозгоспецифическому белку S-100. Исследование проведено в Институте медицинской и биологической кибернетики СО РАМН под руководством академика РАМН М. Б. Штарка совместно с Н. А. Береговым, Н. С. Сорокиной, М. В. Старостиным.
В основу эксперимента был положен известный электрофизиологический феномен длительной посттетанической потенциации, заключающийся в том, что при электрораздражении в тетанизирующем режиме одной из областей среза гиппокампа крыс в другой его области специальным электродом регистрируется суммарный вызванный постсинаптический потенциал. Этот потенциал длительно (от 40 мин до нескольких часов) сохраняется, имеет известные характеристики; его наблюдение позволяет изучать синаптическую эффективность (передачу).
Известно, что формирование длительной посттетанической потенциации - кальций-зависимый процесс. Упомянутый выше белок S-100 является Са-связывающим и принимает важное участие в синаптических процессах, а антисыворотка к белку S-100 эти процессы, в том числе и длительную посттетаническую потенциацию, блокирует.
Однако сочетанное (бипатическое) применение антисыворотки в обычной дозе и ее потенцированной формы СН6 (1(Г12 масс, долей) приводило к отмене указанного синаптического эффекта. Другими словами, гомеопатическая доза вещества (в данном случае - иммунологического препарата) подавляла действие его значимой весовой дозы.
Ценность проведенного эксперимента состоит в том, что в нем использованы физические (электрофизиологические) методы исследований, позволяющие многократно воспроизводить наблюдения в стандартизованных условиях. Для специалистов (нейробиологов, иммунологов), вероятно, небезынтересным будет более подробный отчет о проведенных экспериментах.
В эксперименте было предпринято сопоставление эффектов антител к нейроспецифическому белку S-100, тех же антител в потенцированной форме и, наконец, анализ их сочетанного («бипатического») применения на указанной модели индукции длительной посттетанической потенциации (ДПТП) в гиппокампе.
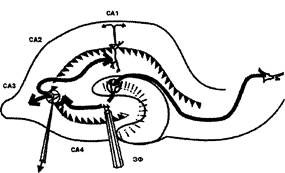
Эксперименты проводили на срезах гиппокампа крыс Вистар (180- 299 г). После приготовления поперечные срезы гиппокампа толщиной 400 мкм помещали в термостатированную камеру (Рис. 13) 35-37°С с проточной средой Ямомото (Рис. 14), аэрируемой карбогеном (95% О2 - 5% СO2). После 40-60 минут инкубации начинали регистрацию вызванных потенциалов, размещая электроды следующим образом: стимулирующий электролитически заточенный биполярный вольфрамовый электрод помещали в область мшистых волокон, регистрирующий стеклянный электрод (толщина кончика - 3-4 мкм, сопротивление - 2-5 Мом, заполненный 2,5 М NaCl) - в область САЗ в зоне начальных сегментов апикальных дендритов (Рис. 15).
* Эксперименты проводились на так называемых переживающих срезах, долго сохраняющих физиологические свойства в искусственной среде.
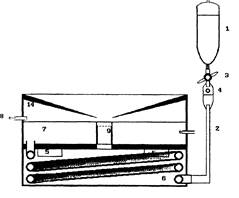
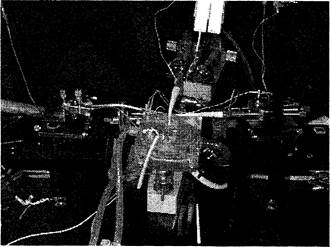
Рис. 14. Общий вид экспериментальной установки для исследований ДПТП в переживающих срезах гиппокампа. Видны камера с проточной системой, микроманипуляторы, несущие катодные повторители, предусилители, раздражающий и регистрирующий электроды. Ориентацию электродов см. на рис. 15.
Рис. 13. Схема экспериментальной камеры для исследования ДПТП на переживающих срезах гиппокампа. Из резервуара (1) среда инкубации по поливиниловым трубочкам (2) через кран (3), позволяющий регулировать скорость потока жидкости и устанавливать ее на уровне 250 мкл/мин, через капельницу (4), чтобы в систему не попадали пузырьки воздуха, подавалась в компартмент (6), содержащий теплоемкую жидкость. Постоянная температура этой жидкости поддерживалась при помощи термостата (5). Среда инкубации, протекая по спиралям трубочки в компартменте (6) нагревалась до нужной температуры и подавалась в камеру (7), объемом 5-10 мл. Отток среды из камеры осуществлялся при помощи водоструйного насоса (8). Инкубационная камера (7) сверху была накрыта крышкой (14). Поскольку жидкость в камере (7) нагревалась и аэрировалась карбогеном, то в ней интенсивно шли процессы испарения воды и обмена углекислым газом между средой и воздухом. Крышка (14) имела единственное отверстие над камерой (9), через которое проникали водяные пары и СО2, увлажняя пространство над камерой и препятствуя дрейфу pH.
Рис. 15. Схема расположения регистрирующего (1) и стимулирующего (2) электродов в связи с исследованием динамики ДПТП в переживающих срезах гиппокампа. CAi-4 - поля аммонова рога гиппокампа; ЗФ - зубчатая фасция.
Тестирование проводили при помощи одиночных прямоугольных стимулов длительностью 200 мкс, наносимых не реже, чем через 5 мин. Амплитуда тестирующих стимулов находилась обычно в диапазоне 10- 30 В. Вызванные потенциалы регистрировали при помощи 12-разрядного АЦП (Digidata, Axon Instruments, Inc., USA) и обрабатывали на ЭВМ, используя пакет программ pClamp-6 (Axon Instruments, Inc.) и Microcal Origin.
Для выработки ДПТП амплитуду тестирующего стимула подбирали таким образом, чтобы величина ответа составляла 50% от максимальной. Тетанизацию проводили тремя последовательными пачками стимулов частотой 200 Гц, длительностью каждой пачки 1 с и интервалом между пачками 2 с. Через 10 мин процедуру тетанизации повторяли. Регистрацию вызванных потенциалов вели не менее 40 мин после первой тетанизации, что позволяло сделать заключение о формировании или отсутствии ДПТП. Критерием выработки потенциации было значительное (в 1,5-2 раза) увеличение амплитуды вызванного постсинаптического потенциала (ВПСП), наблюдавшееся не менее 20 минут после второй тетанизации.
Для анализа влияния антител к белку S-100 использовали следующий протокол эксперимента: на 1-2 срезах каждой партии проводили тетанизацию и дальнейшие эксперименты со срезами этой партии продолжали только в том случае, если наблюдали формирование ДПТП. К среде для инкубации добавляли фиксированные количества антител или контрольных растворов и в течение определенного времени выдерживали в ней срезы. В качестве исходного временного интервала, необходимого для развития эффекта, был использован 20-ти минутный интервал, описанный в работе Левис и Тейлора (Levis D., Teylor Т., 1986), изучавших влияние антисыворотки к нейроспецифическому белку S-100 на индукцию ДПТП в срезах гиппокампа крыс. В дальнейшем время инкубации подбиралось экспериментально. После проведения экспериментов с каждым из разведений экспериментальная камера многократно промывалась дистиллированной водой, этиловым спиртом и полностью высушивалась сжатым воздухом.
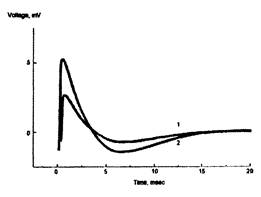
Рис. 16. Пример записи суммарного ВПСП в поле САЗ аммонова рога при внеклеточной регистрации. Амплитуда тестового стимула - 20 В: 1 - до, 2 - через 10 мин после тетанизации. Ось ординат - напряжение в мВ, ось абсцисс - время в мс
На рис. 16 представлена динамика суммарного ВПСП синапсов мшистых волокон зубчатой фасции гиппокампа до и после тетанизации. Формирование ДПТП характеризуется значительным возрастанием амплитуды вызванного потенциала в ответ на тестирующий стимул после проведения тетанизации.
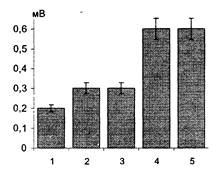
Действие антисыворотки к нейроспецифическому белку S-100 (анти- S-100) в конечном разведении 1:50 приводило после 20 мин инкубации к полному подавлению индукции ДПТП (Рис. 17), как и в ранее проведенных экспериментах. Большая степень разведения антисыворотки, использованная в наших экспериментах, объясняется, вероятно, различиями в титрах антител, полученных в разных лабораториях.
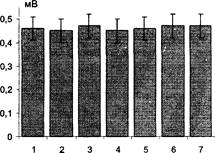
Рис. 17. Влияние AS-100 на выработку ДПТП в срезах гиппокампа. Ось ординат - амплитуда ВПСП (мВ). 1 - до применения AS-100; 2, 3, 4, 5 - в течение 20 мин после применения AS-100 (интервал 5-7 мин); 6, 7 - через 7 и 12 мин соответственно после второй тетанизации. Амплитуда тестового стимула 30 В.
Рис. 18. Развитие ДПТП в присутствии неиммунной сыворотки кролика (в разведении 1:50). По вертикали - средняя амплитуда ВПСП + ошибка среднего. 1 - после 10 минут инкубации в неиммунной сыворотке; 2, 3 - через 5 и 10 минут соответственно после первой тетанизации; 4, 5, 6 - в течение 20 мин после второй тетанизации. Амплитуда тестирующего стимула - 12 В.
Рис. 19. Развитие ДПТП при добавлении 40 мкл 40% этанола. Объем экспериментальной камеры 10 мл. По вертикали - средняя амплитуда ВПСП 1, 2 - при инкубации в среде Ямамото с добавлением этанола в течение 20 мин; 3 - через 10 минут после первой тетанизации; 4, 5 - через 10 и 30 мин соответственно после второй тетанизации Амплитуда тестирующего стимула - 15 В
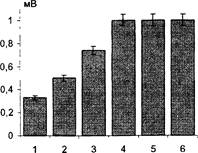
Неиммунная антисыворотка кролика в том же разведении не оказывала влияния на индукцию ДПТП в срезах гиппокампа крыс (Рис. 18). Инкубация срезов с растворами этанола в концентрации, соответствующей таковой в потенцированных препаратах, в том же разведении не приводила к нарушению выработки ДПТП в срезах (Рис. 19).
Для исследования совместного эффекта препаратов использовали сочетанное добавление к среде для инкубации срезов анти-S-l00 (AS-100) и ее потенцированных форм (nAS-100). При этом было установлено, что одновременное добавление указанных веществ не отменяло эффекта анти- S-100, т.е. наблюдалась полная блокада индукции ДПТП в срезах. Аналогичная картина регистрировалась и при 10-минутной преинкубации среза в потенцированной форме анти-S-l00 с последующим добавлением анти-S- 100 и инкубацией среза в растворе обоих веществ еще в течение 20 минут.
20-минутная преинкубация срезов в растворе потенцированной формы анти-S-100 в концентрации 10-12 с последующей 20-минутной инкубацией в растворе обеих (нативной и потенцированной) антисывороток приводила к отмене эффекта анти-S-100, то есть в срезах наблюдали формирование ДПТП, аналогичной таковой в контрольных препаратах, не подвергавшихся воздействию антител (Рис. 20, 21).
Для проверки предположения о том, что причиной отмены действия нативной анти-S-l00 может являться длительная инкубация среза с этанолсодержащим раствором потенцированной формы, приводящая к изменению состояния мембраны и/или нарушению связывания антител с антигеном, была проведена дополнительная серия контрольных экспериментов, в которой преинкубация проводилась в раствором этанола в той же концентрации, что и в потенцированной форме анти-S-l00. Дальнейший же ход эксперимента был аналогичен описанному выше. Как было установлено, при этом сохранялась способность анти-S-l00 блокировать формирование ДПТП.
Рис. 20. Демонстрация «бипатического» эффекта.
А - выработка длительной посттетанической потенциации (ДПТП) в присутствии анти-S- 100 (в конечном разведении 1:50). 1- 3 - при инкубации в среде Ямомото с анти-S-100 в течение 20 мин с интервалами между стимулами 5-7 мин; 4-6 - через 10 мин после 1-й тетанизации с интервалами 3-4 мин; 7, 8 - через 10 и 25 мин соответственно после 2-й тетанизации. Амплитуда тестируемого стимула 12 В.
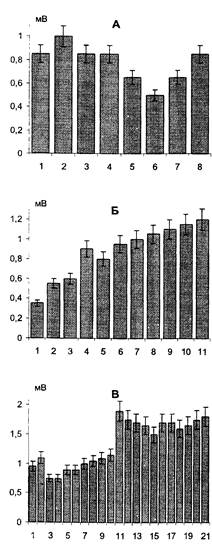
Б - выработка ДПТП в присутствии потенцированной анти-S-100 в концентрации 10-12 (40 мкм). 1-3 - при инкубации в среде Ямамото с nAS- 100 в концентрации 10-12 в течение 20 мин с интервалами между стимулами 5-7 мин, 4-6 - в течение 10 мин после 1-й тетанизации с интервалами 3-4 мин; 7-11 - в течение 30 мин после 2-й тетанизации с интервалами 5-7 мин. Амплитуда тестирующего стимула 20 В.
В - выработка ДПТП в присутствии анти-S-100 в концентрации 10-12 (40 мкл) и в разведении 1:50. 1-3 - при инкубации в среде Ямамото с nAS- 100 в концентрации 10-12 в течение 20 мин с интервалами между стимулами 10 мин; 4-6 - при инкубации с добавлением nAS-100 в течение 20 мин с интервалами 5-7 мин; 7-10 - в течение 10 мин после 1-й тетанизации с интервалами 2-3 мин; 11-21 - в течение 40 мин после 2-й тетанизации с интервалами 3-5 мин. Амплитуда тестирующего стимула 10 В.
Рис. 21 Демонстрация«бипатического» эффекта.
Развитие ДПТП в «бипатическом» режиме. 1,2 - преинкубация с nAS- 100 (10-12) (20 минут); 3, 4 - инкубация с нативной AS-100; 5 - после первой тетанизации; 6, 7 - после второй тетанизации (через 5 и 10 мин соответственно).
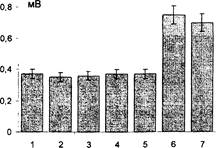
Таким образом, неиммунная сыворотка и этанол в 40% концентрации не препятствовали развитию длительной посттетанической потенциации. Подавление длительной посттетанической потенциации наблюдалось исключительно в растворах антисыворотки к S-100. Преинкубация в потенцированной антисыворотке к S-100 в течение 20 минут снимала эффект подавления длительной посттетанической потенциации антисывороткой к S-100 и не препятствовала нормальному развитию потенцирующей синаптическую эффективность реакции в поле СА3 гиппокампа.
Приведенные в этой главе данные свидетельствуют о реальности бипатического эффекта, проявляющегося в живых организмах, в том числе, в виде феномена системной адаптации.








