 2014-02-05
2014-02-05 10938
10938Эндогенные механизмы регуляции фотосинтеза.
Реализация фотосинтетической функции растения в целом определяется с одной стороны значительной автономностью хлоропластов, а с другой – сложной системой связей фотосинтеза со всеми функциями растения. В ходе онтогенеза в растительном организме всегда присутствуют аттрагирующие зоны (зоны, притягивающие питательные вещества). В аттрагирующих центрах происходит либо новообразование и рост структур, либо интенсивный однонаправленный синтез запасных веществ (клубни, плоды и др.). В обоих случаях состояние аттрагирующих центров определяет величину «запроса» на фотосинтез. Если внешние условия не лимитируют фотосинтез, то ведущая роль принадлежит аттрагирующим центрам. Чем мощнее центры, аттрагирующие ассимиляты, тем интенсивнее фотосинтез.
Второй. важный механизм регуляции фотосинтеза связан с фитогормонами и эндогенными ингибиторами роста и метаболизма. Фитогормоны образуются в разных частях растении, в том числе и хлоропластах, и действуют на процессы фотосинтеза как дистанционно, так и непосредственно на уровне хлоропластов. Дистанционное воздействие осуществляется благодаря регулирующему влиянию фитогормонов на процессы роста и развития, на отложение веществ в запас, на транспорт ассимилятов и т.д., т.е. на формирование и активность аттрагирующих центров. С другой стороны фитогормоны оказывают прямое действие на функциональную активность хлоропластов через изменение состояния мембран, активность ферментов, генерацию трансмембранного потенциала. Доказана также роль фитогормонов, в частности цитокинина, в биогенезе хлоропластов, синтезе хлорофиллов, ферментов ц.Кальвина.
На интенсивность фотосинтеза влияют такие факторы внешней среды, как: интенсивность и качество света, концентрация углекислого газа, температура, водный режим тканей растения, минеральное питание и др.
Интенсивность и спектральный состав света.
Листья высших растений поглощают свет в красной и синей областях спектра – лучи, наиболее эффективные для фотосинтеза.. Отражают листья зеленые лучи. Большая часть (60%) попадающего на лист солнечного излучения не может участвовать в фотохимических процессах, поскольку имеет длину волны, которая не поглощается пигментами листа. Часть света отражается поверхностью листа, рассеивается в виде тепла, тратится на процессы не связанные с фотосинтезом и только 1,5-5% расходуется на фотосинтез (фотосинтетически активная радиация - ФАР).
Зависимость скорости фотосинтеза от интенсивности света имеет форму логарифмической кривой. При низкой освещенности на световой кривой можно выделить точку, когда количество углекислоты, поглощаемой при фотосинтезе и выделяемой при дыхании, равны. Эта точка называется световым компенсационным пунктом (рис.). Увеличение освещенности выше светового компенсационного пункта вызывает постепенное возрастание интенсивности фотосинтеза. При дальнейшем увеличении интенсивности кривая выходит на плато, сто свидетельствует о насыщении процесса связывания углекислоты. В этих условиях процесс фотосинтеза уже лимитируется только содержанием углекислого газа. У светолюбивых видов насыщение происходит при более высокой освещенности (10-40 тыс. люкс), чем у теневыносливых (1000 люкс).
Активность фотосинтеза в области насыщающей интенсивности света лимитилуется концентрацией СО2 и зависит от мощности системы поглощения и восстановления углекислоты. Чем выше способность растения к восстановлению СО2, тем выше проходит световая кривая фотосинтеза
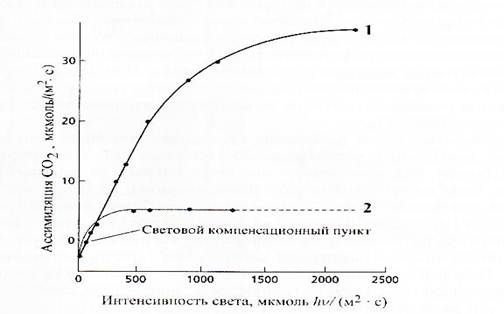
Рис. Изменение интенсивности фотосинтеза у лебеды Atriplex triangularis, выращенной при различной освещенности.
Поэтому у С3-растений насыщение происходит при более низкой освещенности, чем у С4-растений, которые более эффективно связывают углекислоту.
Содержание углекислого газа
Содержание СО2 в атмосфере составляет 0,036%, водяных паров – 2%, кислорода – 21%. азота – более 70%.
СО2 – основной субстрат фотосинтеза. Зависимость фотосинтеза от концентрации углекислоты описывается логарифмической кривой (рис). При концентрации 0,036% интенсивность фотосинтеза составляет лишь 50% и достигает максимума при 0,3%.
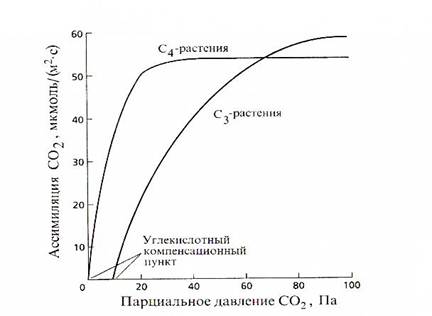
Рис. Зависимость интенсивности фотосинтеза от парциального давления СО2
Многие биологические процессы, в которых участвуют газы (углекислый газ, кислород), определяются не концентрацией, а парциальным давлением. Например, если атмосферное давление 0,1МПа, то парциальное давление углекислого газа составит 36Па (оно вычисляется умножением молярного содержания газа на общее атмосферное давление 0,036х0,1МПа).
У С3-растений при низких концентрациях углекислоты количество СО2 фиксированное при фотосинтезе, меньше чем количество СО2 выделенное при дыхании. При повышении СО2 можно зафиксировать точку, в которой суммарное поглощение углекислоты в фотосинтезе равно 0. Эта концентрация СО2 называется углекислотным компенсационным пунктом. Это параметр характеризует соотношение между процессами фотосинтеза и дыхания в зависимости от содержания СО2 в атмосфере.
Процесс фотосинтеза обычно осуществляется в аэробных условиях. При концентрации кислорода 21%. Увеличение содержания или отсутствие кислорода для фотосинтеза неблагоприятны.
Высокие концентрации кислорода снижают интенсивность фотосинтеза по следующим причинам: 1) повышение парциального давления активирует процесс фотодыхания (РБФ-карбоксилаза ц.Кальвина работает как оксигеназа); 20 кислород окисляет первичные восстановленные продукты фотосинтеза.
Температура
Зависимость интенсивности фотосинтеза от температуры имеет вид параболы с максимумом от 25о-35оС. Однако если концентрация углекислого газа в воздухе будет выше, то температурный оптимум сместится до 35-38оС. Это объясняется тем, что именно при таких температурах активно идут ферментативные реакции (темновая фаза фотосинтеза) (рис.).

Рис. Зависимость интенсивности фотосинтеза от температуры: 1 – при высоком содержании углекислоты; 2 – при 0,036%
Водный режим
Вода непосредственно участвует в фотосинтезе как субстрат окисления и источник кислорода. С другой стороны, величина оводненности тканей определяет степень открывания устьиц и, следовательно, поступления СО2 в лист. При полном насыщении листа водой устьица закрываются, что снижает интенсивность фотосинтеза. Поэтому незначительный водный дефицит благоприятен для фотосинтеза. В условиях засухи происходит закрывание устьиц под влиянием абсцизовой кислоты, которая накапливается в листьях. Длительный водный дефицит приводит к ингибированию нециклического и циклического транспорта электронов и фотофосфорилирования.
Для нормального функционирования фотосинтетического аппарата растение должно быть обеспечено всем комплексом макро- и микроэлементов. Зависимость фотосинтеза от элементов минерального питания определяется их необходимостью для формирования фотосинтетического аппарата (пигментов, компонентов ЭТЦ, структурных и транспортных белков).
Магний входит в состав хлорофиллов, участвует в деятельности сопрягающих белков при синтезе АТР, влияет на активность реакций карбоксилирования и восстановления NADP+.
Железо необходимо для функционирования цитохромов, ферредоксина (компоненты ЭТЦ). Недостаток железа нарушает функционирование циклического и нециклического фотофосфорилирования, синтез пигментов, нарушает структуру хлоропластов.
Марганец и хлор необходимы для фотолиза воды.
Медь входит в состав пластоцианина.
Азот входит в состав хлорофиллов, аминокислот. Недостаток его сказывается активности фотосинтеза в целом.
Фосфор необходим для фотохимических и темновых реакций фотосинтеза. Отрицательно сказываются как недостаток, так и избыток его (нарушается проницаемость мембран)
Калий необходим для формирования гранистой структуры хлоропластов, работы устьиц, поглощения клетками воды. При недостатке калия нарушаются все процессы фотосинтеза.

