 2014-02-05
2014-02-05 6263
6263Режим пастеризации
Созревание молока
Свежевыдоенное молоко – неблагоприятная среда для развития молочнокислых бактерий (вследствие бактерицидных свойств). Для улучшения биологических и технологических свойств молока его подвергают созреванию – выдержке при 10±2°С в течение 14±2 ч. При этом в молоке накапливаются полипептиды, способствующие активизации молочнокислой микрофлоры и повышению кислотности на 1-2°Т. Образующаяся молочная кислота переводит соли кальция из коллоидного в ионизированное состояние, что способствует укрупнению казеиновых частиц:
Са3(PO4)2 ® СаHPO4 ® Са(H2PO4)2
Чем выше кислотность молока, тем быстрее оно свертывается (при повышении кислотности на 1°Т продолжительность свертывания сокращается на 8%) и возрастает скорость синерезиса. При низкой кислотности образуется неплотный вялый сгусток Þ порок резинистая, мажущаяся консистенция. При высокой – сыр крошливой консистенции. Оптимальная кислотность молока для твердых сыров с низкой температурой второго нагревания 17-19°Т, сыров с высокой температурой второго нагревания 18-20°Т, рассольных сыров – 20-21°Т, для мягких сыров – 23-25°Т.
Молоко пастеризуют при низкой температуре 72-76°С с выдержкой 20-25 с. При высокой температуре пастеризации понижается растворимость солей кальция, образуются гидрофильные комплексы b-лактоглобулина с χ-казеином. Þ Увеличивается продолжительность сычужного свертывания молока, образуется дряблый, малосвязный сгусток, увеличивается количество сырной пыли и снижается выход сыра. Для восполнения потерь ионизированного кальция в молоко после пастеризации вносят СаСl2.
Сычужное свертывание молока (сычужная коагуляция) – наиболее важный процесс при производстве сыра.
Более 95% казеина в молоке находится в форме сферических мицелл (частиц, каждая из которых содержит тысячи молекул a-, b-, c-казеинов). Мицеллы казеина состоят из субмицелл. Они удерживаются в составе мицелл коллоидным фосфатом кальция (КФК), который вместе с органическим казеинатом кальция, образует казеинаткальцийфосфатный комплекс (ККФК). Гидрофобные участки a-, b-, c-казеинов погружены внутрь субмицелл, образуя неполярное ядро. Полярные фосфосерильные группы a-, b- казеинов и гидрофильный гликомакропептид (МП) c-казеина окружают ядро, образуя защитный слой и обеспечивая стабильность мицелл в растворе.
Сычужное (энзиматическое) свертывание молока состоит из двух фаз – первичной, или энзиматической, и вторичной, неэнзиматической (коагуляционной). Энзиматическая фаза протекает под действием молокосвертывающих ферментов, коагуляционная фаза идет без участия молокосвертывающих ферментов, но в присутствии ионов Са2+.
Существует несколько теорий, объясняющих механизм сычужного свертывания молока: фосфоамидазная, гидролитическая и др.
Согласно фосфоамидазной теории П.Ф. Дьяченко сычужный фермент, гидролизует белковые фосфоамидные связи без отщепления фосфорной кислоты. При этом в образовавшемся параказеине появляются дополнительные гуанидиновые группы аргинина и гидроксильные группы остатков фосфорной кислоты, которые связывают ионы кальция. Далее между мицеллами параказеина возникают «кальциевые мостики», что приводит к коагуляции белков [32].
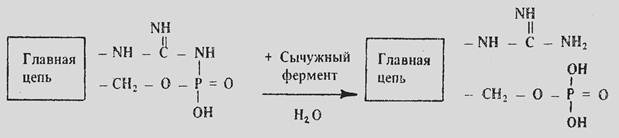
Казеин Параказеин
Предпочтение отдается гидролитической теории.
Энзиматическая фаза
В соответствии с гидролитической теорией под действием молокосвертывающего фермента происходит гидролиз пептидной связи фенилаланин (105) – метионин (106) в полипептидных цепях χ-казеина ККФК, в результате чего молекулы χ-казеина распадаются на гидрофобный пара-χ-казеин и гидрофильный гликомакропептид.
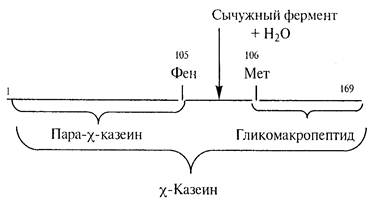
Рисунок 1– Схема ферментативной стадии сычужного свертывания молока
Неэнзиматическая фаза
Отщепление от мицелл гликомакропептида, обладающего высоким отрицательным зарядом и гидрофильными свойствами, приводит к потере устойчивости казеиновых мицелл (снижает заряд мицеллы в два раза, вызывает разрушение большей части гидратной оболочки, ликвидирует ворсистый внешний слой мицелл. В результате, силы электростатического отталкивания между частицами уменьшаются, пространственные факторы стабилизации мицелл ослабевают, что приводит к потере устойчивости казеиновых мицелл). Сущность неэнзиматической фазы состоит в агрегации дестабилизированных мицелл пара-c-казеина за счет сил гидрофобного взаимодействия или посредством «кальциевых мостиков», образующихся в результате присоединения ионов кальция к серинфосфатным группам as- и b-казеина двух или более сблизившихся параказеиновых мицелл [4].
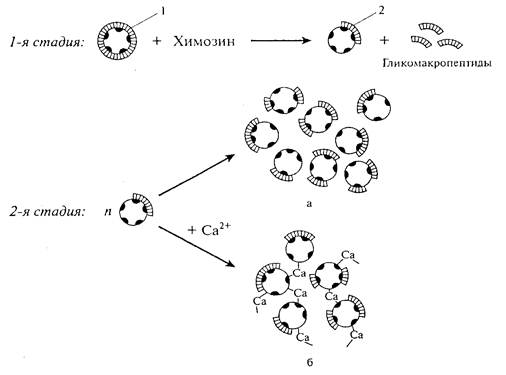
а – коагуляция мицелл под действием сил гидрофобного взаимодействия; б – коагуляция мицелл за счет кальциевых мостиков; 1 – нативные казеиновые мицеллы; 2 – параказеиновые мицеллы
Рисунок 2 – Схема процесса сычужного свертывания молока
При этом мицеллы параказеина собираются в агрегаты, цепочки, которые соединяются продольными и поперечными связями, образуя единую трехмерную сетку – сгусток (происходит гелеобразование). В ячейках сетки заключена дисперсионная среда, содержащая жировые шарики и сыворотку [8].
Таким образом, на стадии свертывания молока параллельно с биохимическим превращением казеинового комплекса в результате действия молокосвертывающих ферментов происходят и физико-химические изменения.
Картину сычужного свертывания молока можно изучить с помощью реологического метода, основанного на измерении эффективной вязкости в процессе свертывания молока.
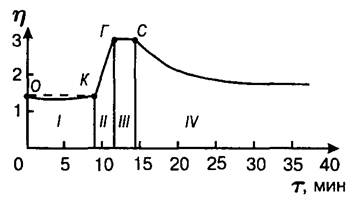
I – индукционный период; II – стадия флокуляции; III – метастабильное равновесие; IV – стадия синерезиса;
О – внесение сычужного фермента; К – начало явной коагуляции; Г – гель-точка; С – начало синерезиса
Рисунок 3 – Реограмма процесса сычужного свертывания молока
По данным ВНИИМС (Табачников, Дудник) процесс сычужного свертывания разделен на четыре стадии:
I – индукционный период (лаг-фаза), включает ферментативную стадию и стадию скрытой коагуляции (участок О-К), заканчивается с началом хлопьеобразования, наблюдаемого визуально;
II – стадия массовой коагуляции (участок К-Г), заканчивается образованием сгустка;
III – стадия структурообразования и упрочнения сгустка (участок Г-С), прекращается с началом синерезиса;
IV – стадия синерезиса, выделение сыворотки из сгустка (после точки С).
Гидролиз c-казеина начинается сразу после внесения в молоко молокосвертывающих энзимов. Визуально, свертывание молока становится заметным после расщепления 80–85% c-казеина (точка К), т.е. вторая (коагуляционная) стадия начинается до окончания первой (энзиматической). Небольшое изменение вязкости происходит во время индукционного периода: после внесения фермента вязкость снижается, что сопровождается небольшим увеличением степени дисперсности казеиновых частиц, к концу индукционного периода вязкость повышается до начальной величины, вследствие агрегации параказеиновых мицелл [10]. На одну молекулу молокосвертывающего фермента, при обычной дозе химозина, приходится около 100 мицелл казеина и необходимо время для их подготовки к флокуляции [10].
После т. К вязкость резко повышается, вследствие массовой агрегации частиц. В гель-точке частицы связываются в пространственную структуру. До гель-точки система сохраняет свойства золя (мицеллы не связаны друг с другом и находятся в свободном броуновском движении), после гель-точки образуется гель – единая пространственная система, обладающая упругими свойствами. При этом вязкость прекращает нарастать. Вязкость на участке Г–С не изменяется. В т. С начинается разделение системы на твердую и жидкую (сыворотка) фазы и вязкость уменьшается.
Участок реограммы О–К соответствует энзиматической стадии (до расщепления 85% казеина), участок К–Г–С – неэнзиматической стадии сычужного свертывания молока. [10]
Факторы, влияющие на характер процесса сычужного свертывания и качество образующегося сгустка: состав и свойства молока, режим пастеризации, активность и состав бактериальной закваски и молокосвертывающего фермента, температура свертывания, доза хлорида кальция и др.

