 2014-02-02
2014-02-02 2675
2675Рельеф
Орографические факторы (характер рельефа, высота над уровнем моря, протяженность, характер склонов и др.) – очень сложно, косвенно действующие факторы. Они не являются экологическими факторами в узком смысле, так как непосредственно на растительные сообщества влияния не оказывают. Тем не менее, роль рельефа как фактора, очень сильно влияющего на климатические и эдафические факторы, чрезвычайно велика. С рельефом связана вертикальная поясность растительного покрова. В большинстве случаев именно разнообразие условий рельефа обеспечивает разнообразие растительных сообществ в пределах одной климатической зоны. Нередко рельеф определяет распространение тех или иных видов растений и, тем самым, видовой состав растительных сообществ.
Принято различать четыре категории рельефа – макрорельеф, мезорельеф, микрорельеф и нанорельеф.
Макрорельеф – это крупные формы рельефа (горные хребты, плоскогорья, равнины, низины), образованные эндогенными процессами и определяющие основные особенности растительности крупного региона. Элементы макрорельефа создают на относительно небольшой площади настолько большую амплитуду высот, что возникает целая серия климатических комплексов. Это связано, в первую очередь с тем, что при подъеме на каждые 100 м температура понижается в среднем на 0,5-0,6 °С. Одновременно с изменением температуры меняются также режим осадков и условия освещения. Все это приводит к тому, что при подъеме в горы можно наблюдать такое явление, как вертикальная поясность. Вертикальная поясность часто схожа с изменениями растительности на равнине, которые можно наблюдать по мере движения от подножия данной горной системы в сторону полюса того же полушария (широтная зональность). Однако эта закономерность не абсолютна, так как в горах можно довольно часто наблюдать выпадение того или иного пояса растительности, инверсию поясов и др. Кроме того, распределение растительности на склонах разной экспозиции даже на одной и той же горе будет очень сильно отличаться.
Помимо формирования вертикальной поясности, макрорельеф также оказывает очень сильное влияние на растительность, преграждая путь ветрам, обладающим определенными свойствами (например, сухим или богатым влагой, теплым или холодным). Такое влияние может распространяться очень далеко за пределы расположения горной системы. Так, преграждающие путь влажным западным ветрам Кордильеры являются причиной того, что на североамериканском континенте природные зоны, начиная с широколиственных лесов, располагаются не с севера на юг, а с востока на запад. Кавказский хребет делает возможным существование в Закавказье субтропиков, препятствуя распространению холодных северных ветров и т.д.
Мезорельеф – средние по размерам формы рельефа с колебаниями высот, как правило, не более нескольких десятков метров (долины малых рек, овраги, второстепенные отроги хребтов, моренные холмы, дюны и др.). Элементы мезорельефа образованы главным образом внешними процессами (деятельность текучих вод, ветра, ледников).
Мезорельеф играет очень важную роль в дифференциации экологических условий в пределах того или иного элемента макрорельефа. Так, при рассмотрении участка равнины, прорезанного рекой, сразу бросается в глаза разница в экологических условиях на плакоре, по склонам коренных берегов реки и непосредственно в речной долине. Северный и южный склоны холма будут очень сильно различаться целым комплексом экологических факторов: температурным режимом, водным режимом, почвами. Зачастую это позволяет сосуществовать на относительно небольшой территории растительным сообществам, представляющим различные зональные типы растительности.
Это свойство мезорельефа позволило В. В. Алехину и Г. Вальтеру в 1951 г. сформулировать правило предварения. Согласно этому правилу, склоны северной экспозиции несут на себе растительные группировки, свойственные более северной растительной зоне или подзоне, а склоны южной экспозиции – растительные группировки, 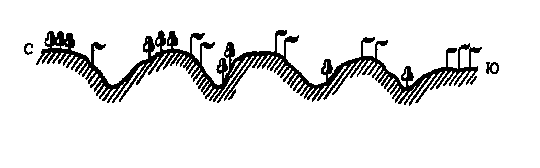 характерные для более южной растительной зоны или подзоны (рис. 14). Это происходит из-за того, что гораздо более сильно прогреваемый склон южной экспозиции будет более благоприятен для формирования более теплолюбивых растительных сообществ, характерных для более южных районов. Склон северной экспозиции, наоборот,
характерные для более южной растительной зоны или подзоны (рис. 14). Это происходит из-за того, что гораздо более сильно прогреваемый склон южной экспозиции будет более благоприятен для формирования более теплолюбивых растительных сообществ, характерных для более южных районов. Склон северной экспозиции, наоборот,
| Рис. 14. Схема правила предварения (Алехин, 1951). |
будет представлять подходящие условия для северных более влаголюбивых растений.
Очень хорошим примером этого правила являются закономерности размещения лесных сообществ в тундровой и степной зонах. Так, в тундре и лесотундре при продвижении с севера на юг участки леса будут появляться лишь на склонах южной экспозиции. В лесной зоне лес является зональным типом растительности и располагается на плакорах. На южной окраине лесной зоны леса сохраняются лишь по склонам северной экспозиции и на дне балок, тогда как по южным склонам произрастает уже типичная степная растительность. Мезорельеф приводит к расширению диапазона экологических условий и, соответственно, к большему разнообразию различных типов местообитаний на определенной территории.
Микрорельеф – мелкие формы рельефа, размеры которых обычно не превышают нескольких метров (степные блюдца, приречные валы, суффозионные западины и др.). Они, как правило, имеют эрозионное или антропогенное происхождение. Наличие микрорельефа приводит к тому, что в пределах того или иного участка местности происходит более тонкая дифференциация экологических условий и формируются различающиеся по составу и строению растительные сообщества. Примером такой дифференциации может служить распределение растительных сообществ в пределах речной поймы.
Речная пойма представляет собой пониженную часть речной долины, которая заливается водой во время паводка. Почвы речной поймы имеют аллювиальное (наносное) происхождение. Весной, во время паводка, вместе с водой река переносит большое количество минеральных и органических частиц, которые ежегодно смываются с плакора и откладываются в пойме. Скорость течения воды во время паводка в разных частях затопленной поймы очень сильно различается: наибольшей она будет в прирусловой части и наименьшей – в притеррасной части поймы. Это приводит к тому, что в прирусловой части поймы происходит отложение лишь самых крупных частиц, переносимых водой. Но, так как таких частиц в любом случае по объему будет больше всего, в прирусловой части поймы за счет песчаных наносов формируется береговой вал – наиболее возвышенная часть поймы. В средней части поймы откладываются более мелкие пылеватые частицы и, наконец, до притеррасной части поймы, там, где течение воды во время паводка наиболее медленное, доносятся лишь самые мелкие илистые частицы. Это приводит к различиям в составе почв: у берега реки почвы наиболее бедные, но наиболее влагопроницаемые, так как состоят главным образом из песка, тогда как вблизи террасы – наиболее плодородные, так как состоят большей частью из мелких органических частиц. Объем наносов, откладываемых рекой во время паводка, также закономерно уменьшается от прирусловой части к притеррасной, что приводит к образованию перепада высот в пойме и, соответственно, разнице в водном режиме почв. Это приводит к тому, что в речной пойме формируется серия растительных сообществ, сильно различающихся своими экологическими условиями, хотя абсолютный перепад высот в пойме реки редко составляет более одного метра.
Нанорельеф – очень мелкие (несколько десятков сантиметров, иногда до 1-2 м по горизонтали и вертикали) формы рельефа, часто имеющие биогенное происхождение. К элементам микрорельефа относятся кочки, кротовины, гниющие стволы поваленных деревьев, крупные камни и др. Нанорельеф приводит к расширению экологической емкости конкретного местообитания и, следовательно, к обогащению его видового состава. Очень часто нанорельеф является одной из причин, приводящих к мозаичности фитоценозов, что будет рассматриваться ниже.
Как говорилось выше, биотические факторы являются следствием взаимоотношений организмов. В результате жизнедеятельности растений, входящих в состав фитоценоза, и связанных с ними гетеротрофных организмов экотоп трансформируется в биотоп. При этом входящие в состав фитоценоза растения, с одной стороны, взаимодействуют друг с другом на уровне так называемых фитогенных полей, а с другой стороны – находятся в сложных многогранных отношениях с гетеротрофными компонентами биоценозов, формируя консорции.
Фитогенное поле означает сферу влияния одного растения на окружающую его среду и соответственно на занимающие эту среду растения. Растение может по-разному воздействовать на среду. Формируя надземные органы (листья, стебли) оно тем самым затеняет нижележащее пространство. Расходуя воду на транспирацию и обеспечение процессов жизнедеятельности, растение иссушает почву. В результате расходования растением элементов минерального питания происходит уменьшение их концентрации или же, наоборот, увеличение за счет разложения поступающего в почву опада и высвобождения извлекаемых корнями растения из более глубоких горизонтов почвы соединений азота, фосфора, калия, кальция и др.
| Рис. 15. Изменение экологической обстановки и напочвенного покрова под елью по профилю ствол (С) – край кроны (КК). По оси абсцисс – расстояние от ствола, по оси ординат – признак. 1 – толщина кроны (по вертикали); 2 – сквозистость кроны; 3 – осадки; 4 – максимальная температура; 5 – опад; 6 – толщина подстилки; 7 – толщина нижней разложившейся части подстилки; 8 – рН подстилки (Ипатов, Кирикова, 1999). |
Радиус фитогенного поля может меняться от нескольких сантиметров у трав, до нескольких метров у крупных деревьев. При этом один и тот же вид в различных частях своего ареала может формировать фитогенные поля различного размера: большие в области эколого-фитоценотического оптимума и меньшие в периферической части ареала вида, там, где условия для него будут пессимальными.
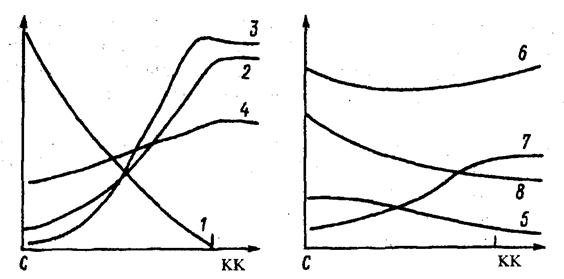 Ярким примером изменений условий среды в пределах фитогенного поля могут служить фитогенные поля единичных особей ели в сосновом лесу. Как видно из рис. 15, при удалении от ствола ели меняется целый комплекс экологических факторов: освещенность, температура, влажность, толщина подстилки, рН почвы и др. При этом специальные опыты показали, что различные факторы по-разному влияют на разные виды: элиминация вереска происходит в результате затенения; брусники, черники, зеленых мхов – дефицита света и влаги, кроме того, зеленые мхи болезненно реагируют на количество опада ели (Ипатов, Кирикова, 1999).
Ярким примером изменений условий среды в пределах фитогенного поля могут служить фитогенные поля единичных особей ели в сосновом лесу. Как видно из рис. 15, при удалении от ствола ели меняется целый комплекс экологических факторов: освещенность, температура, влажность, толщина подстилки, рН почвы и др. При этом специальные опыты показали, что различные факторы по-разному влияют на разные виды: элиминация вереска происходит в результате затенения; брусники, черники, зеленых мхов – дефицита света и влаги, кроме того, зеленые мхи болезненно реагируют на количество опада ели (Ипатов, Кирикова, 1999).
Консорция – это такая совокупность особей различных видов, в центре которой находится особь автотрофного или гетеротрофного вида, компоненты которой связаны с центром трофически, топически, форически или фабрически, и под влиянием которой формируется специфическая микросреда (Голубец, Чернобай, 1983). Консорция является элементарной структурной единицей биоценозов, в которой различные организмы объединены друг с другом. Представление о консорциях в их современном понимании было введено В. Н. Беклемишевым (1951) и Л. Г. Раменским (1952).
Центральная особь консорции называется центральным ядром (детерминантом консорции), а связанные с ней организмы – консортами. В консорции объединяются все трофические группы организмов: автотрофы, фитофаги, зоофаги, сапрофаги, некрофаги, копрофаги, редуценты. Таким образом, консорцию можно рассматривать как структурную единицу биогеоценоза; она имеет свои пространственные и функциональные параметры.
Детерминантом консорции может быть как автотрофный, так и гетеротрофный организм. Соответственно, консорции можно разделить на две группы: автотрофно детерминированные и гетеротрофно детерминированные. Если ядром консорции является отдельная особь, то такие консорции называются индивидуальными. Индивидуальные консорции являются элементарными структурными единицами биогеоценоза. Они, в свою очередь, объединяются в популяционные консорции (совокупность организмов, связанных с популяцией детерминанта консорции), видовые консорции и т. д. Все это не что иное, как объединение (типизация) индивидуальных консорций.
Консорты образуют ряд концентров (рис. 16). В первый концентр входят консорты, непосредственно связанные с детер-минантом консорции или только трофически (животные-фитофаги, сапротрофы, эккрисотрофы), или и трофически и топически (паразитные организмы, симбионты), или только топически (эпифиты, лианы, 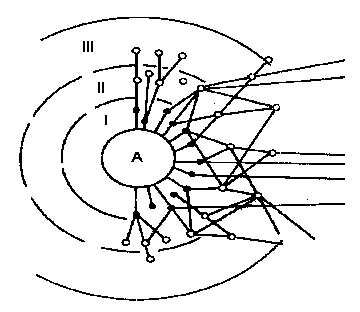 животные, гнездящиеся на детерминантах). Консорты, входящие в состав второго концентра консорций, используют в качестве источника энергии и веществ консортов первого порядка, как в живом, так и в отмершем состоянии, а также экскременты животных-фитофагов и сапрофагов. В соответствии с этим в состав второго
животные, гнездящиеся на детерминантах). Консорты, входящие в состав второго концентра консорций, используют в качестве источника энергии и веществ консортов первого порядка, как в живом, так и в отмершем состоянии, а также экскременты животных-фитофагов и сапрофагов. В соответствии с этим в состав второго
| Рис. 16. Схема консорции (по В. В. Мазингу, 1966): А – детерминант консорции; I, II, III - концентры (Мазинг, 1966). |
концентра входят зоофаги, зоопаразиты, микофаги, бактериофаги, сапротрофы второго порядка, в том числе копротрофы. Аналогична структура последующих концентров консорций.
Рассматривая структуру консорции, не следует забывать о том, что большинство видов второго и третьего концентров не взаимодействуют непосредственно с центральным ядром консорции. Так, во второй и третий концентры консорции растений входит большое количество хищников первого и второго порядка, которые гораздо подвижнее и менее специализированы в пище. Поэтому они объединяют большое количество консорций, тогда как в каждой конкретной консорции они будут являться случайным элементом. Тем не менее, консорты второго и последующих концентров могут оказывать на детерминанта консорции косвенное, нередко значительное влияние, регулируя численность фитофагов и фитопаразитов. Кроме того, стоит отметить тот факт, что некоторые организмы могут одновременно входить в состав нескольких концентров. Так, например, некоторые хищники, являясь организмами второго и третьего концентров, могут поедать плоды растений, то есть входить в состав организмов первого концентра.
Следует подчеркнуть, что консортные отношения нельзя сводить только к цепям питания, поскольку они включают не только трофические, но и топические связи, а также попутное воздействие консортов на детерминанта консорции: выделение метаболитов в их ткани паразитными организмами, опыление цветков, перенос семян и плодов и др.
В состав любой консорции помимо ее детерминанта входит различное число видов-консортов, которое может быть довольно значительным. Так, в состав видовой консорции березы входит 91 вид паразитных и 36 – микоризообразующих грибов, 46 видов эпифитных лишайников, 7 – эпифитных печеночников и 16 – лиственных мхов, 8 видов клещей, 574 – насекомых, 8 – птиц, 9 видов млекопитающих – всего 803 вида, не считая эпифитных водорослей и бактерий (Работнов, 1978). Сюда не включены многочисленные организмы сапротрофного звена. Хотя следует отметить и то, что имеются виды (плауны, хвощи) с довольно малым числом консортов.

