 2014-02-03
2014-02-03 3737
3737МИТОЗ
В основе роста и дифференцировки органов и тканей животных лежит размножение клеток, смена одного клеточного поколения на другое. Клетки тела, или соматические клетки," разных поколений содержат одинаковое количество генетического материала, что обеспечивается особым механизмом деления, получившим название митоз. В процессе митоза выделяют две основные стадии — интерфазу и собственно митоз.
Интерфаза предшествует митозу. В ней выделяют три периода: 1) пресинтетический (Gi); 2) синтетический (S) и 3) постсинтетический (G2). В Gi-периоде в клетке происходит накопление белка, РНК и других продуктов, необходимых для образования клеточных структур и последующего деления. В течение S-периода синтезируется ДНК и происходит ауторепродукция (самоудвоение) хромосом, что приводит к возникновению второй хроматиды. В G2-ne-риоде продолжается синтез ДНК и белков, накапливается энергия.
Время прохождения клетками разных периодов интерфазы неодинаково. Так, по данным Л. В. Трофимовой, клетки костного мозга кур проходят периоды Gi, S, G2 и митоза соответственно за 6, 7, 8 и 23 ч. ДНК в хромосомах также синтезируется с разной скоростью. Например, у коров наибольшей" скоростью репликации характеризуется 14-я хромосома. Задержка в репликации (синтезе) ДНК на отдельных хромосомах может быть причиной возникновения патологии у животных.
Вслед за интерфазой начинается деление клетки — митоз. Выделяют четыре стадии митоза: профазу, метафазу, анафазу, тело-фазу. При изучении митоза основное внимание уделяется поведению хромосом. В профазе хромосомы представляют собой клубок длинных тонких хроматиновых нитей. К концу этой фазы митоза ДЙ|1на их уменьшается за счет спирализации примерно в 25 раз, Шбдюдается также разрушение ядрышка. Предполагают, что ве-Ирство ядрышка участвует в образовании веретена деления. Нити веретена прикрепляются к центриолям, которые в этот период УЖе разделились и находятся на противоположных полюсах клет-Ш. Завершается профаза разрушением ядерной оболочки клетки.
В метафазе утолщенные спирализованные хромосомы перемещаются в экваториальную плоскость клетки. В этот момент Они имеют характерную для каждой из них форму, удобную для Цитогенетического анализа.
Началом анафазы считают момент разделения удвоенных хромосом на хроматиды, которые затем расходятся к противоположным полюсам клетки.
Во время телофазы сестринские хроматиды достигают противоположных полюсов и деспирализуются. Так формируются два ^>черних ядра. Наряду с делением материнского ядра происходит деление цитоплазмы, образование оболочек клеток.
Таким образом, в процессе митоза из одной материнской клетки возникают две дочерние, содержащие такой же набор хромосом, как и у исходной клетки (рис. 5). Основное биологическое значение митоза состоит в точном распределении хромо-
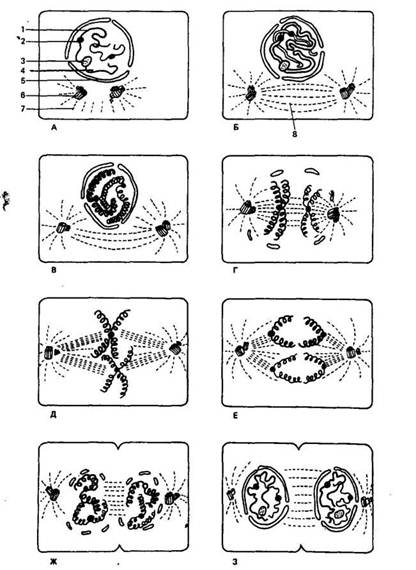
Рис. 5. Схема митотического деления животной клетки (по К. Вилли
и В. Детье, 1975):
А — интерфаза (стадия «покоя»); Б — ранняя профаза (центриоль разделилась); В и Г— более поздняя профаза; Д— метафаза; Е и Ж— ранняя и поздняя анафазы; 3 — телофаза (ядерная оболочка исчезла, началось деление цитоплазмы); / — хромосома; 2 — центромера; 3 — ядрышко; 4— ядро; 5— ядерная оболочка; 6— центриоль; 7— звезда; 8— веретено
сом между двумя дочерними клетками; тем самым сохраняются Преемственность хромосомного набора в ряду клеточных поколения и полноценность генетической информации каждой клетки, что необходимо для осуществления общих и специфических функций живого организма.
Патология митоза. При делении соматических клеток могут возникать различные нарушения, связанные с повреждением хромосом, митотического аппарата, цитоплазмы. К числу этих нарушений относятся задержка митоза в профазе, Нарушения спирализации и деспирализации хромосом, раннее разделение хроматид, фрагментация и пульверизация хромосом, задержка митоза в метафазе и др. Эти нарушения возникают йод действием отдельных химических веществ, радиации, вирусных инфекций. Так, при заражении свиней вирусом чумы наблюдали пульверизацию и фрагментацию (множественные разрывы) хромосом.
Процесс развития половых клеток носит название гаметогене-зв. У самцов этот процесс называется спермиогенезом, а у самок — овогенезом (рис. 6). Половые клетки в развитии последовательно проходят следующие стадии: размножения, роста, созревания и формирования. В стадии размножения клетки интенсивно делятся митотическим путем. В стадии роста клетки накапливают питательные вещества, особенно при овогенезе.
Наиболее ответственный момент с точки зрения генетики при образовании половых клеток — мейоз — процесс редукционного и эквационного деления ооцитов и сперматоцитов, в результате которого образуются половые клетки с гаплоидным набором хромосом. Рассмотрим наиболее важные моменты поведения вдомосом в мейозе. В этом процессе выделяют две стадии деления (см. схему): 1) редукционную и 2) эквационную.
Схема мейоза

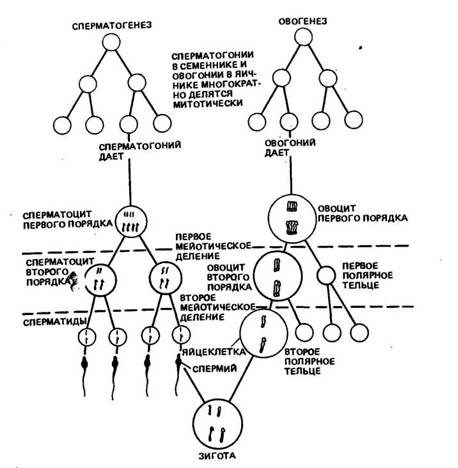
Рис. 6. Сравнение процессов снерматогевеза и овогенеза у животных с гаплоидным числом хромосом, равным 2 (по К. Вилли и В. Детье, 1975)
Непосредственно перед мейозом клетки половых желез находятся в интерфазе.
Редукционное деление начинается с профазы I, которая, как видно из схемы, подразделяется на пять фаз. На первой стадии профазы I — лептонемы хромосомы деспирализо-ваны, они в 2—5 раз длиннее метафазных. Под электронным микроскопом можно видеть, что они состоят из двух хроматид, соединенных центромерой. На следующей стадии — зигонемы наблюдаются притяжение и слияние (конъюгация) гомологичных хромосом. Каждая пара конъюгирующих хромосом образует бивалент, а по числу хроматид — тетраду. На этой стадии происходит образование синаптонемного комплекса (СК), входящего в состав бивалента. Нарушение формирования СК между гомологичными хромосомами наблюдают у гетерозиготных носителей хромосомных аберраций. Далее, на стадии пахинемы, происходят утолщение и укорочение хромосом, так что сестринские хрома-тиды становятся хорошо различимыми; на отдельных из них можно видеть и ядрышки.
Следующая фаза — диплонема характеризуется тем, что конъ-югярующие хромосомы начинают отталкиваться и постепенно расходятся от центромеры к концам. При этом образуются характерные фигуры, напоминающие греческую букву «хи» (х) и получившие вследствие этого название хиазмы. В точках соприкосновения гомологичных хромосом возникают разрывы. Они могут быть одинарными, двойными и более сложными. В результате разрывов образуются фрагменты хроматид, которые затем могут воссоединяться на другой хромосоме, изменяя тем самым комбинацию генетического материала в клетке.
Обмен участками между гомологичными хромосомами получил название кроссинговера.
На последней стадии профазы I — диакинезе происходит резкое укорочение хромосом, так что к концу этой стадии хроматиды остаются связанными только на концевых участках. Этим и заканчивается профаза I. Необходимо отметить, что при более детализированном изучении мейоза в профазе выделяют и другие промежуточные стадии, например пролептонему, диктионему и т. д.
На стадии метафазы I биваленты располагаются в плоскости экватора центромерами к противоположным полюсам. Силы отталкивания здесь увеличиваются.
В анафазе I начинается расхождение гомологичных хромосом к противоположным полюсам, которое носит случайный характер. Каждая из пар гомологичных хромосом имеет одинаковую вероятность распределения в одну из двух дочерних клеток.
В телофазе I хромосомы достигают полюсов клетки. Затем восстанавливаются ядерная оболочка и ядрышко, хромосомы декон-денсируются. В конце телофазы делится цитоплазма (цитокинез) и образуются две дочерние клетки с гаплоидным набором хромосом.
Отличительной особенностью первой стадии мейоза является то, что в период анафазы сами хромосомы не делятся на хроматиды, как при митозе, а лишь расходятся гомологичные пары хромосом к разным полюсам клетки и формируются две дочерние клетки с редуцированным наполовину набором хромосом, состоящим, однако, из двух хроматид.
Между первой и второй стадиями мейоза имеется непродол-жипрелыщй период покоя — интеркинез, во время которого не происходит репродукции хромосом.
Эквационное, или уравнительное, деление аналогично митозу, где клетки последовательно проходят четыре фазы: профазу II, метафазу II, анафазу II, телофазу II. На стадии йнафазы II хромосомы разделяются на две хроматиды, которые ЭДтем с помощью нитей веретена расходятся к противоположным полюсам. На стадии телофазы II заканчивается формирование еще двух клеток. В результате после двух последовательных, стадий мейоза из каждой клетки образуются четыре новые с гаплоидным набором хромосом. Для более наглядного представления всех этих событий можно воспользоваться схемой мейоза, приведенной на рисунке 7.
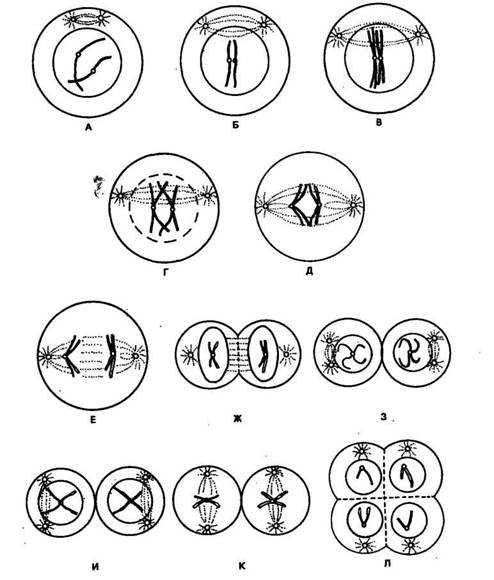
Рис. 7, Схем» последовательных спщнй мейоза (по К. Смясоиу н П. Уэбстеру, 1980):
А — лептонема, предшествующая конъюгации хромосом; Б — начало конъюгации на стадии зигонемы; В— пахинема; Г— диплонема; Д— метафаза I; E— анафаза I;.ЖГ~-телофаза I; 3 — интерфаза между двумя делениями мейоза; Я—профаза II; К— метафаза II; ЛГ—тело-фаза II
Таким образом, в результате двух мейотических делений из одной клетки с диплоидным набором хромосом образуются четыре клетки с гаплоидным набором хромосом и в 2 раза меньшим, чем в соматических клетках, содержанием ДНК. Вероятностный характер распределения материнских и отцовских гомологичных хромосом в разные клетки позволяет создать новые комбинации негомологичных хромосом в яйцеклетках и сперми-ях, чем достигается огромное число новых сочетаний наследственной информации.
Новые сочетания генетической информации возникают вследствие кроссинговера. Каждая из хромосом в метафазе I содержит участки, происходящие от отцовских и материнских хромосом. Рекомбинации хромосом при кроссинговере и вероятностное распределение их по клеткам — причины наследственной изменчивости организма. Мейоз, оплодотворение и митоз обеспечивают поддержание постоянства числа хромосом в смежных поколениях видов. В этом их биологическое значение.
Патология мейоза. Основная патология мейоза — нерасхождение хромосом. Оно может быть первичным, вторичным и третичным. Первичное нерасхождение возникает у особей с нормальным кариотипом. При этом на стадии анафазы I нарушается разделение бивалентов и обе хромосомы из пары гомологов переходят в одну клетку, что приводит к избытку хромосом в данной клетке (л + 1) и недостатку в другой (л—1). Вторичное нерасхождение возникает в гаметах у особей с избытком (трисо-мией) одной хромосомы в кариотипе. В результате этого в процессе мейоза образуются и биваленты, и униваленты. Третичные нерасхождения наблюдают у особей, имеющих структурные перестройки хромосом, например транслокации. Нерасхождение хромосом отрицательно влияет на жизнеспособность организма животных. Подробно об этом будет изложено в последующих главах.
Отличительная особенность мейоза у самок — образование в первом и втором мейотических делениях так называемых полярных телец, которые впоследствии дегенерируют и в размножении не участвуют. Неравные деления в овогенезе обеспечивают яйцеклетке необходимое количество цитоплазмы и запасного желтка, чтобы она могла выжить после оплодотворения. В отличие от спермиогенеза, который у самцов происходит как во внутриутробный (пренатальный) период, так и после рождения (постна-тальный период), у самок яйцеклетка после рождения не образуется. Однако к концу пренатального периода у самок накапливается огромное количество овоцитов (у коров, например, десятки тысяч), а созревают и дают начало развитию потомства лишь немногие из них. Этот резерв генетического материала, особенно у малоплодных животных, таких, как крупный рогатый скот, в настоящее время начинают использовать для искусственного стимулирования созревания многих яйцеклеток, последующего их оплодотворения и пересадки (трансплантации) специально подготовленным коровам-реципиентам. Таким образом от одной ценной коровы в год можно получить не одного, а несколько телят.
Оплодотворение наступает после слияния гаплоидных сперматозоида и яйцеклетки и образования диплоидной клетки — зиготы, дающей начало развитию эмбриона. При делении клеток эмбриона, содержащих хромосомы матери и отца, генетическая информация поступает во все клетки нового организма.
Морфологические и функциональные различия хромосомных наборов — основные причины эмбриональной смертности после оплодотворения при скрещивании разных видов или бесплодия гибридов. Так, при скрещивании зайца и кролика не происходит имплантации и развития оплодотворенной яйцеклетки. Эмбриональная смертность наступает при скрещивании козы и овцы. Мужские гибщады осла (2л = 62) и лошади (2л = 64), как известно, не &щ т потомства, они стерильные, или бесплодные. Стерильность гибридов-самцов наблюдается при скрещивании европейского крупного рогатого скота с бизоном, зубром, яком, а также гауром и гаялом.
Это связано с тем, что у гибридов нарушаются процессы спермиогенеза. Однако при скрещивании европейского крупного рогатого скота с азиатским горбатым зебу, дикого кабана (2л = 36) и домашней свиньи (2л = 38) потомство рождается плодовитое. В результате сложных вариантов скрещиваний в последнее время удалось получить плодовитых гибридов крупного рогатого скота с зуС^ом, бизоном, яком. Гибриды отличаются такими ценными качествами, как крепкое телосложение, высокая жизнеспособность, хорошие мясные признаки.
Контрольные вопросы.
1. Каковы морфологическое строение и химический состав хромосом?
2. Что такое кариотип и каковы его особенности у разных видов животных?
3. Что такое митоз н каково его биологическое значение?
4. Что такое гаметогенез и каковы его особенности у самцов и самок?
5. Что вы знаете о мейозе и его биологическом значении?
6. Каковы основные формы патологий митоза и мейоза?

