 2014-02-18
2014-02-18 9172
9172Липиды представляют большую группу разнообразных веществ, содержащихся в животных и растительных тканях.
Общие признаки липидов.
1. Плохая растворимость в воде и хорошая растворимость в органических растворителях (эфире, бензоле, ацетоне).
2. По строению большинство липидов являются сложными эфирами.
Функции липидов.
1. Липиды наряду с глюкозой являются источником энергии для организма.
2. Многие липиды являются биологически активными веществами – гормоны, витамины, ненасыщенные жирные кислоты, простаглаандины.
3. Липиды являются обязательным компонентом клеточных мембран.
   Липиды Липиды |  Производные липидов Производные липидов | ||
 Простые (дикомпонентные) Простые (дикомпонентные) |   Сложные (поликомонентные) Сложные (поликомонентные) | 1. Каротины 2. Стериды | |
| Триглицериды (жиры) | Фосфолипиды | Гликолипиды | 3. Жирорастворимые витамины |
Важным компонентом липидов являются карбоновые кислоты с длинными углеводородными радикалами.
Жирные кислоты делят на:
1. Насыщенные или предельные
2. Ненасыщенные или непредельные
Ненасыщенные жирные кислоты делят на:
1. Мононенасыщенные (содержащие 1 двойную связь)
2. Полиненасыщенные – несколько двойных связей.
Наибольшее значение имеют полиненасыщенные жирные кислоты, содержащие 2 и более кратных связей.
| Жирные кислоты | |
| Насыщенные | Ненасыщенные |
| Мононенасыщенные | Полиненасыщенные |
 3 3 | 6 |
- номер атома углерода, от которого начинается двойная связь, начиная от -углерода.
Нумерация углеродных атомов в жирной кислоте начинается с карбоксильной группы. Углеродный атом метильной группы на дальнем (дистальном) конце назван -углеродом. Положение двойной связи представлено знаком 3 с номером в верхнем индексе. Индекс обозначает положение двойной связи между 3 и 4 атомами углерода, начиная с -углерода.
В настоящее время для обозначения положения двойной связи используют (омега). - это моследний, удаленный атом углерода, а 3 и 6 обозначают положение двойной связи.
3 и 6 – жирные кислоты обладают разными свойствами и функциями.
1. Насыщенные жирные кислоты в основном энергетический материал (используются и как структурный материал).
2. Полинасыщенные жирные кислоты – эссенциальные соединения. Они не синтезируются в организме (линолевая) или синтезируются в недостаточном количестве.
3. 3 – необходимы для построения клеточных мембран.
4. 6-необходимы для синтеза регуляторных молекул: простагландинов, лейкотриенов, тромбоксанов.
В клетках человека наиболее часто встречаются следующие жирные кислоты:
| Предельные: | ||
| 1. Пальмитиновая | С15Н31СООН | 16: 0 |
| 2. Стеариновая | С17Н35СООН | 18:0 |
| Непредельные: (ненасыщенные) | ||
| 1. Олеиновая | С17Н33СООН | 18:1; 9 9 |
| 2. Линолевая | С17Н31СООН | 18:2; 9, 12 6 |
| 3. Линоленовая | С17Н29СООН | 18:3; 6, 9, 12, 6 |
| 4.Арахидоновая | С19Н39СООН | 20:4, 5, 8, 11, 14, 6 |
Ненасыщенные жирные кислоты подразделяются на моно- и полиненасыщенные. Наибольшее значение имеет арахидоновая кислота и ее производные. Из арахидоновой кислоты синтезируются простагландины, простациклины и лейкотриены, тромбоксаны.
Простогландины (ПГ) содержат циклопентановое кольцо, с которым связаны две боковые цепи. В зависимости от числа двойных связей в боковых цепях различают ПГ1, ПГ2, ПГ3. В зависимости от наличия кето или гидроксогруппы простогландины обозначают как А,В и так далее.
Простогландины – это короткодействующие регуляторы многих процессов (сокращения гладких мышц, кровообращения, передачи нервных импульсов, поддержания водного и электролитного баланса).
В тромбоцитах из арахидоновой кислоты образуются тромбоксаны, которые вызывают агрегацию тромбоцитов и ускоряют процесс свертывания крови.
В лейкоцитах из арахидоновой кислоты синтезируются лейкотриены. Лейкотриены являются мощными активаторами сокращения гладких мышц.
Триглицериды – это сложные эфиры глицерина и высших жирных кислот. Триглицериды делят на простые и сложные. Простые триглицериды содержат 3 одинаковых остатка жирных кислот, а сложные триглицериды содержат остатки разных жирных кислот.
СН2ОСOR СН2ОСOR1
| |
CHOCOR СНОСOR2
| |
СН2ОСOR СН2ОСOR3
В природных жирах преобладают жирные кислоты с четным числом углеродных атомов (С16, С18), с неразветвленной углеродной цепью.
К сложным липидам относятся фосфолипиды. В состав фосфолипидов входят глицерин, два остатка высших жирных кислот, остаток фосфорной кислоты и азотсодержащие соединения:
СН2ОСOR1
|
СНОСOR2 O Кефалин
| //
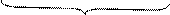 СН2 – О – P – O – CH2 – CH2 – NH2
СН2 – О – P – O – CH2 – CH2 – NH2
| Коламин (этаноламин)
OH
СН2ОСOR1
|
СН2ОСOR2 O Лецитин
| //
СН2 – О – P – O – CH2 – CH2 – N(СН3)3
 |
|
OH холин
СН2ОСOR1
|
СН2ОСOR2 O Серинфосфатиды
| //
СН2 – О – P – O – CH2 – CH2 – СООН
| |
 OH NH2
OH NH2
серин
 Фосфолипиды – дифильные соединения (есть полярная и неполярная часть).Фосфолипиды всегда расположены на поверхности раздела. Вода – неводный сегмент (клеточные мембраны).
Фосфолипиды – дифильные соединения (есть полярная и неполярная часть).Фосфолипиды всегда расположены на поверхности раздела. Вода – неводный сегмент (клеточные мембраны).
 - гидрофильная часть (глицерин)
- гидрофильная часть (глицерин)
- гидрофобные хвосты Ж.К.
R2 в ФЛ всегда полиненасыщенная, чаще всего 3
Производные липидов
Стериды – производные высокомолекулярных полициклических ненасыщенных спиртов. Представителем стеридов является холестерин и холестериды.
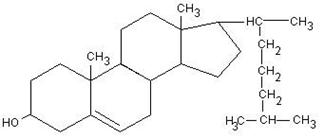 Холестерин и холестериды гидрофобны.
Холестерин и холестериды гидрофобны.
Холестерин входит в состав клеточных мембран, он необходим растущему организму.
Каротины синтезируются в растительных клетках, в основном  -каротины. Чем больше каротинов в пище и крови, тем реже возникают злокачественные новообразования. Производные -каротинов является витамин А.
-каротины. Чем больше каротинов в пище и крови, тем реже возникают злокачественные новообразования. Производные -каротинов является витамин А.
Жирорастворимые витамины А, Е, К. Сейчас витамин D относят к гормонам.
Транспорт липидов
Транспорт липидов осуществляется липопротеидами.
Липопротеид состоит из гидрофобного ядра, образованного триглицеридами и холестерином. Снаружи гидрофобное ядро окружено белком, это фактор стабильности липопротеида (заряд белковой молекулы и водная оболочка не позволяет частицам слипаться).
Фосфолипиды связаны с белком, а хвост фосфолипида погружен в гидрофобное ядро. Фосфолипиды выполняют функцию связывания за счет дифильности. Липопротеид – это не макромолекула, т.к. здесь есть гидрофобные взаимодействия, слабые полярные взаимодействия. Это надмолекулярная структура, она может терять или приобретать новые компоненты.
1. Хиломикроны – самые крупные липопротеиды. Имеют низкую плотность (d 0,94 г/см3).Содержат 2% белка, 98% липиды, в основном триглицериды, которые поступают с пищей.
0,94 г/см3).Содержат 2% белка, 98% липиды, в основном триглицериды, которые поступают с пищей.
Синтезируются в кишечнике, переносят в основном триглицериды пищи, холестерин из кишечника в жировую ткань и печень. Триглицериды в составе хиломикронов легко гидролизуются в капиллярах жировой ткани и других периферических тканях.
В составе липопротеидов обязательно есть белок. Белки липопротеидов называются аполипопротеинами. Они бывают А,В,С,Д,Е (апо А, апо Е).
Хиломикроны содержат апо-А,В,С,Е.
Плазма крови желтая, прочная, аполесцирующая жидкость. После еды плазма мутная, хилезная, из-за высокого содержания хиломикронов.
2. Липопротеиды очень низкой плотности – ЛПОНП (ЛОНП), плотность (0,94 – 1,006). Содержат 5 – 7% белка, остальные липиды (ТГ + холестерин). ЛПОНП транспортируют эндогенные триглицериды, которые синтезировались в печени, к жировой ткани. В крови ЛПОНП, обогащаясь холестерином превращаются в ЛНП.
Аполипопротеиды представлены апо В,С,Е.
3. Липопротеиды низкой плотности (ЛПНП = ЛНП) – содержат 10 – 15% белка, 50% холестерина. Основная транспортная форма холестерина и холестеридов. Образуется в плазме крови и обеспечивает перенос холестерина к периферическим тканям. Плотность = 1,006 – 1,063 ЛНП – апо-В
4. Липопротеиды высокой плотности (ЛПВП = ЛВП), плотность 1,063 – 1,210. 50% белка, 50% распределены между липидами. Уносят избыток холестерина от тканей к печени. Образуются в крови. ЛВП – апо-Д.
Клинициста интересуют ЛНП и ЛВП.
5. Альбумин + НЭЖК – транспортная форма неэстерифицированных жирных кислот.
Функции аполипопротеинов:
Аполипопротеины В легко связываются с рецептором к ЛНП и проникают в клетку.
Обмен триглицеридов.
В полости рта ТГ не подвергаются расщеплению из-за отсутствия ферментов. В желудке взрослых людей ТГ также не расщепляются, активность липазы желудочного сока низкая.
У детей в желудке, где рН желудочного сока около 5, происходит переваривание эмульгированного жира молока.

 Переваривание ТГ начинается в 12-перстной кишке, после их эмульгирования желчными кислотами.
Переваривание ТГ начинается в 12-перстной кишке, после их эмульгирования желчными кислотами.
Гидролиз эмульгированных ТГ ступенчатый ферментативный процесс. Сначала гидролизуются сложноэфирные связи в положении 1 и 3, а затем 2.
Продукты гидролиза всасываются: глицерин диффундирует в клетку эпителия кишечника, жирные кислоты с короткой углеродной цепью всасываются путем диффузии, а с длинной углеродной цепью в виде холеиновых кислот. Холеиновые кислоты – это комплекс НЭЖК+2-4 молекулы желчных кислот. В слизистой кишечника комплексы распадаются, жирные кислоты по системе воротной вены идут в печень, а из глицерины и НЭЖК синтезируются эндогенные триглисериды.
Обмен триглицеридов.
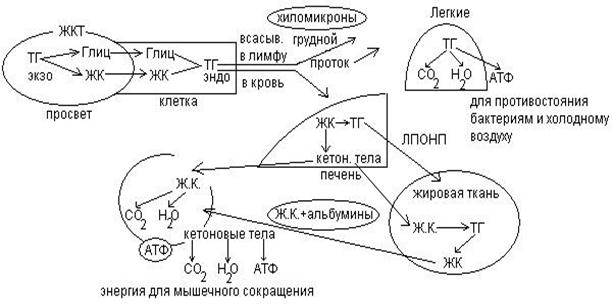
Транспортные формы липидов.
1. хиломикроны – крупные, рыхлые, 2% белка, 98% ТГ пищи.
Плазма крови – желтая, позрачная, опалесцирующая жидкость. После еды плазма мутная, хилезная. Так будет пока хиломикроны не будут утилизированы.
2. ЛПОНП – 5-7% белка, остальное ТГ+холестерин, но это эндогенные триглицериды, которые синтезировались в печени.
3. ЛПНП – 10-15% белка, 50% холестерина. Это основная транспортная форма холестерина и холестеридов.
4. ЛПВП – 50% белка, остальные 50% распределены поровну между липидами.
ЛПНП приносят холестерин к тканям.
ЛПВП уносят избыток холестерина от тканей к печени.
5. Альбумины+НЭЖК
Белок липопротеидов – аполипротеин: А, В, С, D, Е.
Хиломикроны А, В, С, Е
ЛПОНП В, С, Е
ЛПНП В
ЛПВП D
Апо В – легко связывается с рецептором и ЛПНП проникает в клетку.
Обмен триглицеридов.
Липиды – разнообразные по химическому строению вещества, нерастворимые в воде и хорошо растворяющиеся в органических растворителях.
Значение липидов в организме:
1. Липиды являются одним из компонентов клеточных мембран.
2. Липиды являются источником энергии для организма.
3. Липиды входят в состав водоотталкивающих и термоизоляционных покровов.
Наиболее распространенные липиды – это нейтральные жиры или триацилглицеролы, или триацилглицериды.
Триацилглицериды – это сложные эфиры, образованные трехатомным спиртом глицерином и высшими жирными кислотами.
Природные жирные кислоты содержат четное число атомов углерода и имеют неразветвленную цепь. В состав триглециридов входят следующие предельные жирные кислоты: пальмитиновая – С15Н31СООН, стеариновая – С17Н35СООН и непредельные жирные кислоты – олеиновая – С17Н33СООН (содержит одну двойную связь), С17Н31СООН – линолевая (содержит две двойных связи), С17Н29СООН – линоленовая кислота (содержит три двойные связи).
Переваривание и всасывание липидов
В полости рта жиры не подвергаются расщеплению, так как слюна не содержит ферментов, расщепляющих жиры. У взрослыхлюдей жиры проходят через желудок без изменений, так как липаза желудочного сока малоактивна.
Переваривание жиров в полости желудка играет важную роль у детей. рН желудочного сока у детей около 5,0, что способствует перевариванию эмульгированного жира молока.
Расщепление жиров пищи происходит в верхних отделах тонкого кишечника, где есть и условия для эмульгирования жиров. Наиболее мощное эмульгирующее действие на жиры оказывают соли желчных кислот.
Гидролиз эмульгированных триглицеридов под действием панкреатической липазы происходит постадийно сначала быстро гидролизуются сложноэфирные связи 1 и 3, а потом идет гидролиз 2-моноглицерида.
Основными продуктами, образующимися в кишечнике, являются жирные кислоты, моноглицериды и глицерин.
Всасывание жиров происходит в проксимальной части тонкого кишечника. Тонкоэмульгированные жиры частично могут всасываться через стенку кишечника без предварительного гидролиза. Однако основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные кислоты, моноглицериды и глицерин. Жирные кислоты с короткой углеродной цепью и глицерин свободно всасываются в кишечнике и поступают в кровь воротной вены, оттуда – в печень. В составе мицелл высшие жирные кислоты и моноглицериды переносятся от места гидролиза жиров к всасывающей поверхности кишечного эпителия.
Ресинтез жиров в стенке кишечника
В стенке кишечника синтезируются жиры специфичные для организма и отличающиеся по строению от пищевого жира.
Механизм ресинтеза тригицеридов в клетках кишечника сводится к следующему: первоначально из жирных кислот образуется ацил-КоА, после чего происходит ацилирование моноглицеридов с образование ди-, а затем триглицеридов.
Однако в эпителиальных клетках кишечного эпителия содетжатся ферменты – моноглицеридлипаза, расщепляющая моноглицериды на глицерин и жирную кислоту, и глицеролкиназа, способная превращать глицерин в глицерол-3-фосфат. Глицерол-3-фосфат взаимодействует с активной формой жирной кислоты – ацил-КоА, образуя фосфатидную кислоту, которая используется для ресинтеза тригицеридов.
Промежуточный обмен липидов.
Метаболизм липидов включает следующие основные процессы:
1. Расщепление триглицеридов в тканях с образованием высших жирных кислот и глицерина – липолиз.
2. Мобилизацию жирных кислот из жирового депо и их окисление.
4. Биосинтез жирных кислот.
5. Биосинтез триглицеридов.
6. Биосинтез фосфоглицеридов.
7. Биосинтез холестерина.
Внутриклеточный липолиз
Главным эндогенным источником жирных кислот служит резервный жир, содержащийся в жировой ткани.
Так как в качестве источников энергии могут использоваться только свободные жирные кислоты, то триглицериды сначала гидролизируются под действием липаз – до глицерина и свободных жирных кислот.
Свободные жирные кислоты из жировых депо могут переходить в плазму крови, после чего они используются тканями и органами в качестве энергетического материала.
В жировой ткани содержится несколько липаз: триглицеридлипаза, диглицеридлипаза и моноглицеридлипаза. В результате липолиза образуются глцерин и свободные жирные кислоты, которые с током крови попадают в органы и ткани, где комплекс распадается, а жирные кислоты подвергаются -окислению частично, а также используются для синтеза триглицеридов, фосфолипидов, этерификации холестерина.
В 1904 г. Франц Кноп пришел к выводу, что жирные кислоты расщепляются путем окисления при -углеродном атоме.
В 1949 г. Юджин Кеннеди и Альберт Ленинджер обнаружили, что окисление жирных кислот происходит в митохондриях. Перед проникновением в митохондрии жирные кислоты подвергаются активации.
1. Активация жирной кислоты происходит в митохондриальной мембране, где она катализируется ацил-КоА-синтетазой:
R – СООН + АТФ + НSКоА  R – СОSКоА + АМФ + РР1
R – СОSКоА + АМФ + РР1
В результате этой реакции образуется Акцил-КоА и одновременно АТФ расщепляется до АМФ и неорганический пирофосфат. Это сопряженная реакция: энергия, высвобождающаяся при расщеплении АТФ на АМФ и пирофосфат, используется в активном центре фермента для образования новой тиоэфирной связи.
2. Перенос остатка жирной кислоты через мембрану митохондрий осуществляется карнитином:
Ацил-КоА + карнитин  Ацил-карнитин + НSКоА
Ацил-карнитин + НSКоА
На внутренней мембране митохондрий происходит регенерация КоА-производных жирных кислот и Ацил-КоА поступает в матрикс митохондрий:
Ацилкарнитин + НSКоА Ацил-КоА + карнитин
В процессе действуют два пула КоА – цитозольный и митохондриальный. Эти пулы выполняют разные функции. Митохондриальный пул КоА используется для окислительного расщепления пирувата, жирных кислот и некоторых аминокислот, тогда ка цитозольный пул участвует в биосинтезе жирных кислот.
Процесс окисления жирных кислот в митохондриях состоит из 2 стадий. На первой стадии происходит отщепление двухуглеродных фрагментов – в виде ацетил-КоА.
На второй стадии окисления жирных кислот остатки ацетил-КоА окисляются в цикле лимонной кислоты до СО2 и Н2О. Таким образом, ацетил-КоА, образующийся в результате окисления жирных кислот, поступает на общий конечный путь окисления вместе с ацетил-КоА, образующимся из глюкозы через реакцию окисления пирувата.
На обеих стадиях окисления жирных кислот атомы водорода или соответствующие им электроны передаются по митохондриальной цепи переноса электронов на кислород. С этим потоком электронов сопряжен процесс окислительного фосфорилирования.
Этапы - окисления
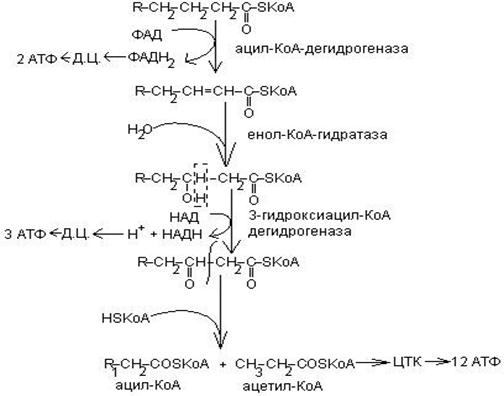
Атомы водорода, отщепляемые от ацил-КоА, переносятся на ФАД, затем на убихинон и дальше по дыхательной цепи к кислороду. В результате переноса этой пары протонов и
ацил-КоА, переносятся на ФАД, затем на убихинон и дальше по дыхательной цепи к кислороду. В результате переноса этой пары протонов и  по дыхательной цепи к кислороду образуются две молекулы АТФ путем окислительного фосфорилирования АДФ.
по дыхательной цепи к кислороду образуются две молекулы АТФ путем окислительного фосфорилирования АДФ.
На третьем этапе окисления жирных кислот имеет место передача протонов и на НАД. Образовавшийся в этой реакции НАДН передает затем восстановительные эквиваленты НАДН – дегидрогеназе дыхательной цепи. На каждую пару электронов, переходящих по цепи переноса электронов от НАДН к кислороду образуется 3 молекулы АТФ.
Расчет выхода энергии при окислении жирной кислоты
1. Подсчет количества молекул Ацетил-КоА, образовавшихся при окислении жирной кислоты:
N = 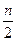
п – число атомов углерода в молекуле жирной кислоты
2– число атомов углерода в молекуле Ацетил-КоА
2. Число витков - окисления
- 1 (число витков -окисления на единицу меньше, чем число молекул СН3CОSКоА, т.к. при окислении четырех углеродного остатка жирной кислоты сразу образуется 2 молекулы СН3СОSКоА)
3. Количество АТФ, образующихся в процессе -окисления
1 виток -окисления - 5 АТФ
(- 1) витков -окисления - х АТФ
4. Количество АТФ, образующихся при окислении N молекул Ацетил-КоА в цикле лимонной кислоты.
1 молекула СН3СОSКоА - 12 АТФ
N молекул СН3СОSКоА - y
5. Общее количество АТФ
5 (число витков -окисления) + 12 (число молекул СН3СОSКоА) = 1 АТФ (затрата на активацию жирной кислоты) = выход АТФ при окислении жирной кислоты.
Окисление ненасыщенных жирных кислот
Окисление ненасыщенных жирных кислот в принципе происходит так же, как и окисление насыщенных жирных кислот. Однако здесь имеются некоторые особенности. Двойные связи природных ненасыщенных жирных кислот (олеиновой, линолевой) имеют цис-конфигурацию, а в КоА-эфирах ненасыщенных жирных кислот, двойные связи имеют транс-конфигурацию. Кроме того, последовательное удаление двууглеродных фрагментов при окислении ненасыщенных жирных кислот до первой двойной связи дает 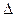 3,4-ацил-КоА, а не 2,3-Ацил-КоА, который является промежуточным продуктом при -окислении насыщенных жирных кислот.
3,4-ацил-КоА, а не 2,3-Ацил-КоА, который является промежуточным продуктом при -окислении насыщенных жирных кислот.
В тканях существуют ферменты, которые осуществляют перемещение двойной связи из положения 3,4 в положение – 2,3, а также изменяют конфигурацию двойной связи из цис-положения в транс-положение:
СН3-(СН2)7 – СН = СН – (СН2)7 – СООН + АТФ + НSКоА
олеиновая кислота 
СН3 – (СН2)7 – СН = СН – (СН2)7 – СОSКоА + АМФ + ФФн
оленоил – КоА -окисление
3СН3 – СОSКоА + СН3 – (СН2)7 – СН = СН – СН2 – СОSКоА
3,4 – цис – еноил – КоА
СН3 – (СН2)7 – СН2 – СН = СН – СОSКоА
-окисление 2,3 – транс – еноил – КоА
6СН3 – СОSКоА
Окисление жирных кислот с нечетным числом углеродных атомов
Из жирных кислот с нечетным числом атомов углерода образуется при -окислении пропионил – КоА. Кроме того, пропионил – КоА образуется при распаде некоторых аминокислот (валина, изолейцина, треонина, метионина).
Пропионил – КоА окисляется по особому пути:
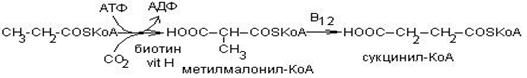
Вначале происходит карбоксилирование с образованием метилмалонил – КоА. В состав фермента входит витамин Н – биотин. Затем метилмалонил – КоА под действием метилмалонилмутозы, содержащей витамин В12, превращается в сукцинил – КоА, который является метаболитом цикла Кребса.
При недостатке витамина В12 эта реакция замедляется и с мочой выводятся большие количества метилмалоната и пропионата.
Биосинтез жирных кислот
Жирные кислоты синтезируются из ацетил-КоА. Несмотря на то, что все реакции -окисления обратимы, этот путь не используется для синтеза жирных кислот.
Основным местом синтеза жирных кислот является цитозоль. -окисление происходит в митохондриях.
На первом этапе происходит превращение ацетил – КоА в малонил – КоА. Этот процесс протекает при участии ацетил – КоА – карбоксилазы, которая содержит витамин Н или биотин. Главную роль в синтезе жирных кислот выполняет пальмитилсинтетаза, которая обеспечивает удлинение углеродной цепи за счет присоединения ацетильного и малонильного остатков к SH – группам активного центра фермента.
Схема синтеза
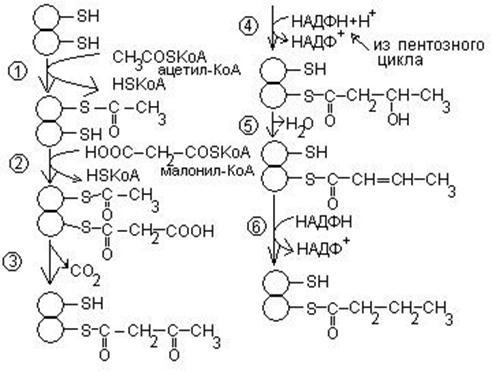
Процесс повторяется до образования пальмитиновой кислоты.
СН3СОSКоФ+СО2+АТФ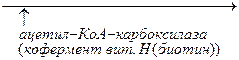
НООС-СН2-СОSКоА+АДФ
Малонил-КоА
Механизм переноса ацетил – КоА через мембрану
Перенос ацетил – КоА через мембрану осуществляется в виде цитрата, который образуется при участии щук. В цитоплазме цитрат распадается на ацетил – КоА и щук.
| Митохондрии | Цитоплазма |
Ацетил-КоА+ЩУК  цитрат цитрат | ЩУК+Ацетил-КоА цитрат |
Пальмитиновая кислота служит предшественником всех жирных кислот. Удлинение углеродной цепи происходит за счет дополнительного присоединения ацетил – КоА или малонил – КоА.
Синтез ненасыщенных жирных кислот
Большинство непредельных жирных кислот образуются путем дегидрирования предельных кислот. Линолевая кислота не синтезируется в организме и должна поступать с пищей.
Наиболее интенсивно синтез жирных кислот происходит в печени, жировой ткани, молочных железах.
Для синтеза жирных кислот необходим НАДФ Н2, который образуется в пентозном цикле.

