 2014-02-18
2014-02-18 963
963Метаболические основы электрогенеза. Метаболизм медиаторов в норме и при патологии. Роль антиоксидантов, антигипоксантов, протекторов мембран.
В этом подразделе мы будем исходить из того, что читатель уже имеет представление об основах нейрофизиологии и нейроанатомии. Поэтому мы обсудим группу заболеваний, в механизме развития которых отчетливо просматриваются биохимические аспекты: миастению гравис, инсульт, заболевания, развившиеся вследствие мутаций в митохондриальной ДНК, синдром ломкой Х-хромосомы и другую патологию, обусловленную повтором триплетов в ДНК, болезнь Паркинсона, болезнь Альцгеймера и шизофрению.
Все названные нервно-психические расстройства характеризуются хроническим течением и нарушением интеллектуальных функций, приводящим к деградации личности. Примечательные данные приводит Национальный фонд исследования мозга (США). Только в этой стране прямые затраты на заболевания, связанные с расстройством функций мозга (психиатрические, неврологические, алкоголизм и др.), составляют более 401 млрд долларов в год или 1/7 всех расходов на здравоохранение США.
Для понимания развития миастении гравис необходимо разобраться в механизме нервно-мышечной передачи
Для этого заболевания характерны периодические приступы мышечной слабости. В заболевание преимущественно вовлекаются мышцы, которые иннервируются черепно-мозговыми нервами. Улучшение наступает после приема лекарственных препаратов, ингибирующих ацетилхолинэстеразу. При миастении гравис, по непонятным причинам, в организме образуются аутоантитела против ацетилхолиновых рецепторов, локализованных в нервно-мышечных синапсах. Эти антитела вызывают разрушение рецепторов и, таким образом, приводят к заметному уменьшению их количества. Чтобы понять последствия такого явления, необходимо знать основные принципы проведения нервного импульса в мышцу.
Функционирование нервно-мышечных синапсов и их блокада
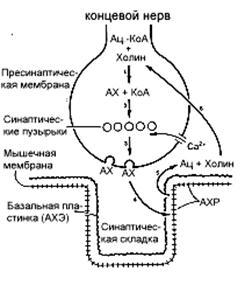
Рисунок 18.7. Схематическое изображение некоторых событий, происходящих в нервно-мышечном синапсе
Часть нервного окончания расположена в тесном соседстве с мышечной концевой пластинкой. Весь процесс, посредством которого АХ стимулирует свой рецептор, можно разделить на 6 этапов (см. текст). Ац - ацетил; КоА - кофермент А; АХ - ацетилхолин; АХЭ - ацетилхолинэстераза; АХР - ацетилхолиновый рецептор.
На рис.18.7 схематически изображен нервно-мышечный синапс и основные события, которые там происходят. Синапс состоит из нервного окончания, отделенного от постсинаптического пространства синаптической щелью. В синапс вовлечена также специальная часть мембраны миоцита (мышечного волокна) - моторная концевая пластинка. В синаптических ямках в непосредственной близи к нервному окончанию содержится большое количество ацетилхолиновых рецепторов (АХР). Каждый нервно-мышечный синапс содержит несколько миллионов рецепторов. По меньшей мере, 400 тысяч из них обеспечивают ток ионов, достаточный для деполяризации концевой пластинки, развития потенциала действия и последующего сокращения мышцы.
Весь процесс проведения нервного импульса в мышцу можно разделить на 6 этапов (рис.18.7). (1) В цитозоле нервного окончания синтезируется ацетилхолин (АХ). Реакцию взаимодействия ацетил-КоА с холином с образованием АХ и КоА-SH катализирует фермент - холинацетилтрансфераза или ацетилхолинэстераза (АХЭ). (2) Затем АХ включается в малые мембраносвязанные частицы, получившие название "синаптические пузырьки". (3) На следующем этапе происходит высвобождение АХ из синптических пузырьков в синаптическую щель. Это происходит путем экзоцитоза, который включает слияние пузырьков с пресинаптическими мембранами. Этот процесс в достаточной степени комплексный (рис.18.8). Импульс достигает нервного окончания и деполяризует его плазматическую мембрану. Деполяризация приводит к непродолжительному открытию потенциалозависимых Са2+-каналов в этой мембране. Поскольку концентрация Са2+ вне клетки более чем в 1000 раз превышает концентрацию свободного Са2+ внутри её, он устремляется внутрь нервного окончания. Растущая концентрация Са2+ в его цитозоле и приводит к высвобождению ацетилхолина в синаптическую щель.
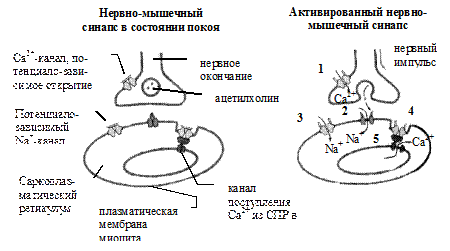
Рис.18.8. Система ионных каналов в нервно-мышечном синапсе
В состоянии покоя, таким образом, спонтанно и единовременно высвобождается около 10000 молекул медиатора (предположительно, содержимое 1 синаптического пузырька). В результате на концевой пластинке формируются небольшие по величине потенциалы. Когда в процессе передачи нервного импульса происходит деполяризация нервного окончания, открываются потенциало-чувствительные Са2+ каналы, через которые Са2+ из синаптического пространства проникает в нервное окончание. Этот Са2+ играет важную роль в процессе экзоцитоза и высвобождения АХ в синаптическое пространство (высвобождается, приблизительно, содержимое 200 пузырьков).
(4) Высвободившийся АХ быстро диффундирует через синаптическую щель к рецепторам, локализованным в синаптических складках. Связывание двух молекул АХ с рецептором приводит к конформационной перестройке последнего. Тогда в рецепторе открывается специальный канал, с помощью которого через мембрану проникают катионы. Поступление ионов Na+ приводит к деполяризации мышечной мембраны и формированию потенциала концевой пластинки. Это, в свою очередь, обусловливает деполяризацию прилежащих мышечных мембран и генерацию потенциалов действия. Последний распространяется на все мышечное волокно, и оно сокращается. (5) Когда канал закрывается, АХ подвергается гидролизу под влиянием АХЭ. Происходит следующая реакция:








