 2014-02-18
2014-02-18 650
650В покоящейся мышце места связывания для миозина на актине замаскированы, и миозин находится в высокоэнергетическом конформационном состоянии (М*). Энергия гидролиза АТФ необходима, чтобы перевести миозин из низкоэнергетического (М) в высокоэнергетическое состояние. Это иллюстрирует уравнение 1:
(М-АТФ) «(М*-АДФ-Фн) Уравнение 1
Когда в цитозоле увеличивается концентрация кальция, и места связывания миозина на актине демаскируются, - формируется актомиозиновый комплекс. Впоследствии от комплекса последовательно отщепляются Фн и АДФ, а миозин переходит в низкоэнергетическое состояние (уравнение 2,3). При этом происходит перемещение соответствующей тонкой нити к М-линии саркомера.
(М*-АДФ-Фн) + А «(М*-АДФ-А) + Фн Уравнение 2
(М*-АДФ-А) «(М-А) + АДФ Уравнение 3
Этапы, приведенные в уравнениях 2 и 3, составляют молекулярный механизм движущей силы мышечного сокращения. Ещё раз подчеркнем, что источником энергии для этой движущей силы является АТФ. Гидролиз нуклеотида опосредует превращение низкоэнергетического конформационного состояния миозина в высокоэнергетическое. Актомиозиновый комплекс существует до тех пор, пока не происходит связывание АТФ. Присоединение АТФ к миозину - это экзергоническая реакция, результатом которой является отход головки миозина от актина (уравнение 4). То есть, АТФ необходим для мышечного расслабления, а в расслабленной мышце миозин находится в высокоэнергетическом конформационном состоянии. То, что конечный продукт уравнения 4 (М-АТФ) является первым участником уравнения 1, свидетельствует о том, что именно в этом месте завершается цикл сокращения.
(МА) + АТФ «(М-АТФ) + А Уравнение 4
Схематическое изображение реакций, приведенных в уравнениях 1-4, в том виде, в котором они происходят в мышцах, показано на рис.7.11.
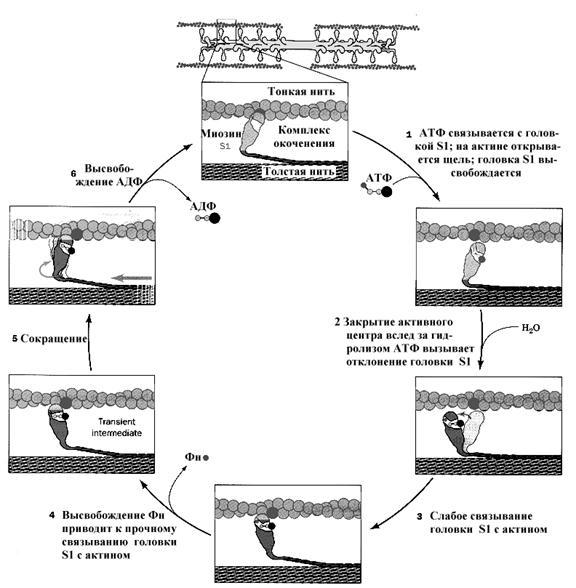
Рис. 17.11. Гидролиз АТФ управляет работой цикла ассоциации-диссоциации актина и миозина








