 2014-02-18
2014-02-18 795
795Несмотря на то, что модель скольжения толстых и тонких нитей достаточно адекватно отражает механизм сокращения во всех мышечных группах, имеются значительные различия между поперечно-полосатой и гладкой мускулатурой. Это вытекает хотя бы из того факта, в гладких мышцах потеря тропонина не приводит к утрате сократительной способности, она продолжает регулироваться уровнем цитозольного кальция. Объяснение пришло позже, когда установили, что в регуляцию перемещения тропомиозина гладких мышц к - и от мест связывания миозина на тонких нитях вовлечен белок, связывающий Са2+/кальмодулин. Белок назвали кальдесмон. Было установлено, что он участвует в регуляции перемещения тропомиозина гладких мышц к - и от центров связывания миозина на тонких нитях.
В общих чертах, картина выглядит следующим образом. Увеличение уровня цитозольного кальция приводит к повышенному его связыванию с кальмодулином. В свою очередь, комплекс кальция с кальмодулином связывает кальдесмон. Тем самым кальдесмон отщепляется от своего места связывания на тонких нитях. Одновременно наблюдается изменение местоположения тропомиозина в спиральной бороздке F-актина и активируется актомиозиновая АТФ-аза. Когда количество кальция уменьшается, комплекс его с кальмодулином диссоциирует; кальдесмон отщепляется от этого комплекса и соединяется с тонкими нитями. Соответственно, ингибируется активность актомиозиновой АТФ-азы. В сущности, кальдесмон здесь выполняет функцию тропонина в качестве зависимого от кальция (Са2+/кальмодулина) регулятора местонахождения тропомиозина на тонких нитях.
Другой отличительной особенностью гладких мышц является наличие в составе миозина уникальной короткоцепочечной полипептидной цепи, получившей название «легкая цепь Р». Эта субъединица может находиться в фосфорилированном и дефосфорилированном состоянии. Присоединение к ней остатка фосфорной кислоты катализирует специальный фермент – киназа легкой цепи миозина, которая также зависит от комплекса Са2+/кальмодулин. В отсутствие этого комплекса гладкие мышцы неподвижны, киназа неактивна, а легкие Р-цепи нефосфорилированы. Повышение концентрации этого комплекса приводит к активации киназы легких цепей миозина, соответственно, к фосфорилированию легких Р-цепей и сокращению. Дело в том, что для сокращения гладких мышц необходимы фосфорилирование легких Р-цепей миозина и отщепление кальдесмона от тонких нитей. Цепь вышеперечисленных событий, причастных к сокращению гладких мышц, приведена на рис.17.12.
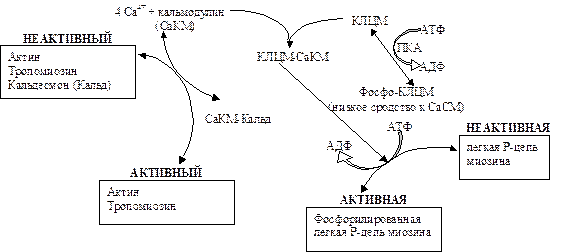
Рис.17.12. Молекулярный механизм сокращения гладких мышц
Увеличение концентрации Са2+ до 10-5 моля вызывает образование комплекса кальция с кальмодулином (СаКМ), который активирует тонкие нити за счет связывания кальдесмона (Кальд) и освобождая места связывания миозина на тонких нитях. СаКМ также связывает и активирует киназу легкой цепи миозина (КЛЦП). Активная киназа катализирует фосфорилирование легкой цепи-Р миозина, в результате активируется актомиозиновая АТФ-азная активность головок миозина. Связывание адреналина с b-адренорецепторами увеличивает количество цАМФ и, следовательно, активность зависимой от цАМФ протеинкиназы (ПКА). В результате снижается сродство МЛЦК к СаКМ, тем самым регулируется сила сокращений, вызванных увеличенным уровнем цитозольного кальция.








