 2014-02-12
2014-02-12 2405
2405Компартментализация нуклеоид (ДНК ядро (ДНК отделена от
и организация не отделена цитоплазмы ядерной
генетического от цитоплазмы оболочкой),
материала содержащей более одной хромосомы,
мембраной), деление ядра путем
состоящей митоза в ядра и некото-
из одной хромо- рых органеллах.
сомы; митоз.
Локализация ДНК в нуклеотиде и в ядре и в некоторых
плазмидах не органеллах
ограниченных
элементарной
мембраной
Цитоплазматические отсутствуют имеются
Органеллы
Рибосомы в 70S-типа 80S-типа
цитоплазме
| Движение цитоплазмы | отсутствует | часто обнаруживается |
| Клеточная стенка (если имеется) | в большинстве случаев содержит пептидогликан | пептидогликан отсутствует |
| жгутики | Нить жгутика построена из белковых субъединиц | Каждый жгутик содержит набор микротрубочек собранных в группы |
Таким образом, отметив отличительные признаки прокариот и эукариот, следует подчеркнуть общие свойства микроорганизмов (кроме подчеркнутых выше общих черт строения клетки). Малая величина особи. Она не только послужила причиной отделения этих организмов от животных и растений: с нею также связаны особенности морфологии микробов, активность и пластичность их метаболизма и распространение их в природе, а также удобство обращения с ними в лаборатории. Как было показано выше, большое отношение поверхности к объему приводит к более интенсивному взаимодействию с внешней средой; с этим связан очень быстрый обмен веществами между средой и клеткой многих микроорганизмов. Правило Рубнера (1893) гласит, что энергетический обмен животного в покое пропорционален не массе, а поверхности его тела. Если это правило, в соответствии с его смыслом, распространить на отдельные ткани и малые клетки, то следует ожидать, что уровни метаболической активности будут различаться на несколько порядков. Экспериментально показано, что интенсивность метаболизма, измеренная по потреблению О2, действительно зависит от размеров клеток и тканей. Соответственно высоки и скорости прироста микроорганизмов, для примера: в организме быка весом 500 кг за 24 часа прирост белка составляет 0,5 кг; за это же время 500 кг дрожжей могут синтезировать более 50000 кг белка. 2. Пластичность метаболизма. Из за небольших размеров клетки у микроорганизмов содержится лишь несколько сотен тысяч белковых молекул, т. е. не нужные в данный момент ферменты не могут храниться в такой клетке про запас. Некоторые ферменты, служащие для переработки питательных веществ, синтезируются клеткой микроорганизма только тогда, когда соответствующее вещество появляется вблизи клетки. Такие ферменты носят название индуцибельиых и могут составлять до 10 % общего белка клетки. Набор постоянных ферментов клетки называется конститутивным. Именно этим объясняется высокая способность к адаптации у микроорганизмов, в отличие от высших растений и животных, у которых обмен веществ довольно жестко ограничен имеющимся набором ферментов. Таким образом, следует подчеркнуть, что клеточные регуляторные механизмы у микробов играют существенно большую роль и проявляются более отчетливо, чем у других живых существ. 3. Распространение микроорганизмов. Бактерии вездесущи, в природных условиях, в естественной среде они входят в любой биоценоз, легко разносятся воздушными потоками. В естественной среде нет необходимости в специальном заражении каким-либо микробом. Этим свойством пользуются микробиологи для получения накопительных культур, достаточно взять 1 г садовой земли, чтобы найти вид бактерий, способный расти за счет любого природного вещества. Т. е., если создать условия можно накопить достаточное количество бактерий для дальнейших исследований, выделения чистой культуры. Микроорганизмы существуют повсюду; среда определяет лишь то, какие формы будут в данном месте активно размножаться, создавая в пробирке соответствующие селективные условия, можно из небольшого количества земли, ила, других сред получать накопительные культуры, а из них и чистые культуры большинства известных микроорганизмов. Микроорганизмы принимают активное участие в круговороте веществ в природе, в очистке природных биоценозов от ксенобиотиков и поллютантов, т. е. в процессах очистки окружающей среды. Количественные работы и успехи генетических исследований. Методы, с помощью которых можно выращивать в лаборатории микроорганизмы, разработали О. Брефельд, Р. Кох и его школа в прошлом веке. Введение в практику прозрачных питательных сред, уплотненных желатиной или агаром, позволило изолировать отдельные клетки, следить за их ростом в колонии и получать чистые культуры. Разработка стандартных методов стерилизации и приготовления питательных сред привела к быстрому развитию медицинской микробиологии. Хотя еще Кох описал количественные методы, их преимущества при работе с микроорганизмами были поняты только в последние 50 лет. Малые размеры микроорганизмов позволяют получать в одной пробирке или чашке Петри и исследовать популяции, состоящие из 10 отдельных клеток, и благодаря этому выявлять такие редкие события, как мутация или передача приобретенного признака, не нуждаясь в сложных вспомогательных средствах и довольствуясь малым пространством. Огромные успехи биохимических и генетических исследований не в последнюю очередь достигнуты благодаря легкости обращения с бактериями. Микроорганизмы на службе человека. Конечно в первую очередь мы видим вред приносимый микроорганизмами, патогенными микроорганизмами и их специфическими особенностями занимаются медицинская и ветеринарная микробиология, а также фитопатология. В настоящее время преобладающей является полезная роль микроорганизмов – это, во-первых, классические микробиологические производства. Из дешевых, богатых углеводами отходов путем брожения, осуществляемого клостридиями и бациллами, можно получать ацетон, бутан ол, 2-пропанол, бутандиол и др. важные химсоединения. Производство антибиотиков. Микроорганизмы используются химиками в качестве катализаторов для осуществления некоторых этапов в длинной цепочке реакций синтеза; м/б процессы по своей химической специфичности и по выходу продукта превосходят химические реакции; ферменты: амилазы, протеиназы; пектиназы; «облагораживание» сырья, успехи генной инженерии. По словам Л. Пастера «Нет прикладных наук, но каждая наука имеет много практических приложений».
ТЕМА 2.
ВОЗНИКНОВЕНИЕ И РАЗВИТИЕ МИКРОБИОЛОГИИ
Давно, несколько тысячелетий назад, когда третья планета солнечной системы получила нового обитателя – «единственное существо, сознающее свое место в природе».Линней в 18 веке в своем сочинении «Система природы» назвал Homo sapiens (человек разумный).Человек постепенно знакомился с окружающей его средой, вживаясь в нее, стараясь использовать ее в своих целях. Так уж устроен человек, что стремится все использовать себе на пользу, стремиться расширить свои познания. Человек начал развивать скотоводство, т.е. знакомиться с животным миром; стал землепаш-цем за счет освоения мира растений. Но кроме растений и животных были в окружающей среде и другие существа, которые сопровождали человека от его рождения до смерти. О них он ничего не мог знать, потому что не мог их видеть. В процессе покорения природы человек использовал этих невидимых спутников даже не догадываясь об их существовании. Человек научился печь хлеб из кислого теста, делать вино из перебродившего сока плодов, готовить пиво из зерен различных сортов, получать кисломолочные продукты: кумыс, уксус при силосовании кормов, мочке льна и т.д. В течении последующих веков человек все глубже и шире познавал и осваивал природу: он стал удобрять почву, изменять течение рек, покрывать луга и поля каналами, осваивать моря и океаны, наконец подниматься высоко в горы. Но всегда и повсюду человека сопровождали невидимые существа: одни из них поражали его частыми и неожиданными болезнями, а такие как чума, холера, тиф, оспа оставили в истории человечества страшные страницы, как и детские болезни, такие как дифтерия например, возбудителем которой является палочка Леффлера. Врача, который ценой собственной жизни спасшего больного ребенка. Процессы, вызываемые микробами, люди знали и использовали с незапамятных времен. Издавна они умели готовить виноградное вино, квас, кумыс, кислое молоко, сыр и другие продукты.
Более 2000 лет назад в Индии и Китае владели искусством предохранения от некоторых болезней. В то время прививки против оспы проводились примерно так же, как в настоящее время. Прививочным материалом служили растертые стенки пустул, в которых содержался возбудитель заболевания.
Уильям Гарвей, ученый описавший систему кровообращения человека, впервые в истории подверг сомнению теорию самозарождения организмов и высказал известное «Все живое из яйца» (Omne vivum ex ovo), т.е. при данном состоянии природы живые организмы никогда не возникают из неживой материи, а всегда от себе подобных. После открытия микробов встал вопрос: есть у микробов родители или они появляются из неживой материи? Опыт английского натуралиста Джона Нидхема, вскипятившего бараний бульон, который он поместил в сосуд и плотно заткнул пробкой. Через несколько дней в бульоне появились бактерии. Вновь заговорили о теории самозарождения, т.е. бактерии появляются из неживой материи. Конечно же, в этой мертвой материи заключена сила, способствующая зарождению живых «зверушек». Однако, противники теории самозарождения, итальянский ученый Лаззаро Спалланцани, французский физик Каньяр де ля Тур (изучал брожение пива, видел дрожжи, которые и вызывают брожение, эти мелкие организмы, быстро размножаются в бродящей жидкости и никогда не возникают из неживой материи. Процесс брожения идет только в их присутствии и ими же самими и вызывается. Уже в 19 веке и немецкий естествоиспытатель Теодор Шванн (доказал, что мельчайшие микробы, обнаруживаемые в мясе и являются причиной его гниения), опровергли теорию самозарождения окончательно. Спалланцани повторил опыты Нидхема и установил, что в хорошо прокипяченном бульоне пропадает «жизненная сила», бактерии не появляются. Однако, стоит открыть сосуд с бульоном и с воздухом поступают бактерии и в бульоне их становится много, т.е. при продолжительном кипячении погибают все бактерии и они вновь поступают в него с воздухом. Кроме того, под микроскопом он наблюдал деление бактерий и таким образом он выступил с утверждением, что бактерии происходят от себе подобных. На этом, грубо говоря, закончился первый период развития микробиологии, который можно назвать ознакомительным. Во второй половине XVII в. при бурном развитии торговли и мореплавания возникла необходимость в изготовлении подзорных труб и увеличительных стекол для них. Люди стали овладевать искусством шлифования. Как полагают многие исследователи, первым шлифовальщиком стекол был профессор римского колледжа Афанасий Кирхер (1601–1680). Микроскоп Кирхера – это разрисованная картонная трубочка длиной около 2 см, с одного конца которой вставлена двояковыпуклая линза, с другого – простое плоское стекло с прикрепленными к нему объектами исследования. С помощью такого увеличительного устройства удалось рассмотреть строение блох и червей в мясе. Более мелкие детали исследователь не видел. «Блошиные стекла», как их называл Кирхер, давали увеличение, едва превышающее десятикратное.
Подзорные трубы для флота начали готовить в некоторых морских странах, одной из которых была Голландия. В этой стране (г. Делфт) жил натуралист (по профессии торговец) Антоны ван Левенгук (1632–1723). Он не имел специальной подготовки, но был человеком редких способностей: выдувал стеклянные изделия и обрабатывал металл.
Все свое свободное время он посвящал шлифованию маленьких, но мощных линз. Пользуясь этими линзами, Левенгук часами рассматривал различные предметы: пробку, листья растений, слюну, соскобы с зубов и др. Во многих субстратах он обнаружил живые организмы, которых назвал «анималькулями» – зверьками. Впоследствии им был сконструирован прибор с увеличением до 300 раз, напоминающий микроскоп. Левенгук настолько заинтересовался своими исследованиями, что начиная с 1674 г. стал посылать подробные отчеты о них Королевскому обществу в Лондоне, прилагая рисунки и срезы птичьего пера, сердцевины бузины, пробки. Он описал также красные кровяные тельца (эритроциты), гладкие и поперечнополосатые мышцы, сперматозоиды, инфузории, стебли одно- и двудольных растений и многое другое. Таких писем было около 200. В одном из посланий он писал: «Я видел в материале множество простейших животных, весьма оживленно двигавшихся. В моем рту их больше, чем людей в Соединенном Королевстве». Рассматривая различные материалы, Левенгук обнаружил палочковидные, шаровидные, извитые и другие формы микробов. Это было началом развития морфологического периода в микробиологии. Свои наблюдения Левенгук описал в книге под названием «Тайны природы, открытые Антони ван Левенгуком» (1695).
После подтверждения сделанных им открытий видными в то время учеными Робертом Хуком и Нехеми Грю, А. Левенгук получил признание и славу. В 1698 г, в г. Делфте его посетил русский царь Петр Великий, который не только распорядился купить микроскоп, но и выписал из Голландии опытного мастера по шлифованию оптических стекол. Позднее в Академии наук начали изготовлять первые отечественные микроскопы.
Первым исследователем «анималькулей»м – микроскопических организмов в России был врач-микробиолог М. М. Тереховский (1740–796). В работе «О наливочном хаосе Линнея» он экспериментально отверг теорию о самопроизвольном зарождении жизни. В то время многие исследователи занимались только описанием форм и размеров микробов. Начало физиологическому периоду в микробиологии положено работами французского ученого-химика Луи Пастера (1822–1895).
Во времена Пастера большое количество вин во Франции портилось и страна несла колоссальные убытки. Пастер, изучив болезни вина, установил, что в нем развивается много посторонней микрофлоры (плесень, дрожжи и др.). Он считал, что посторонняя микрофлора попадает из воздуха и с используемой аппаратуры. Для предотвращения болезней вина ученый предложил прогревать его в течение нескольких минут при 50—60 °С, в результате чего некоторые микроорганизмы погибали и продукт не портился при транспортировании. Такой метод предохранения продуктов от порчи получил имя автора и стал называться пастеризацией.
В дальнейшем Пастер показал, что брожение и гниение вызываются также микроорганизмами. Если прогреть бродящие или гниющие субстраты, микробы гибнут, прекращаются вызываемые ими процессы. Таким образом, было доказано, что начало жизни дают невидимые простым глазом организмы.
Изучая процессы маслянокислого брожения, Пастер открыл микроорганизмы, которые могут жить без доступа кислорода воздуха — анаэробы. Своими исследованиями он указал хирургам пути борьбы с микробами при операциях.
Будучи профессором Высшей нормальной школы (Париж), Пастер изучал болезнь шелковичных червей– пебрину. Он установил инфекционную природу болезни и разработал меры профилактики, что явилось «прелюдией» к изучению инфекционных болезней. Используя метод аттенуации (ослабления микробов), Луи Пастер приготовил вакцины против холеры кур, сибирской язвы и других инфекций. Эффективность препаратов против сибирской язвы он продемонстрировал публично в Пуйи-ле-Форе (Франция). Для опыта было взято 60 голов рогатого скота. Животные находились в разных помещениях. Первая группа (24 овцы, одна коза и 6 коров) была вакцинирована 5 и 17 мая 1881 г. соответственно I и II вакцинами Пастера. 31 мая все вакцинированные и невакцинированные животные (24 овцы, одна коза и 4 коровы) были объединены и заражены вирулентной культурой возбудителя сибирской язвы. Через 48 ч (2 июня) было установлено, что вакцинированные животные остались живы, а невакцинированные почти все (за исключением коров, у которых образовались отеки и повысилась температура тела) погибли. Этим опытом Луи Пастер доказал эффективность полученной вакцины.
Последним неоценимым даром, который Пастер сделал человечеству, было предупреждение от бешенства. Для приготовления вакцины против болезни, возбудитель которой не виден в световом микроскопе, надо было обладать гениальной интуицией – предсказать, что неизвестный агент находится в мозговой ткани, сократить инкубационный период на кроликах, ослабить возбудителя и только потом использовать его для предупреждения болезни! Причем приготовленную мозговую суспензию вводить не один, а несколько раз.
Пастер не был врачом, поэтому испытание прививок на людях было для него большим риском. Тем более что такие верные его помощники и друзья, как Эмиль Ру и Шарль Шамберлан, отказались делать первую прививку больному, которого укусила бешеная собака. Они считали, что пока не будет отработана методика прививок на собаках, людей лечить нельзя. Однако эмоции и настоятельные просьбы родственников больного возобладали над разумом. 6 июля 1885 г. Пастер отважился спасти жизнь сильно искусанному бешеной собакой девятилетнему эльзасскому мальчику Жозефу Мейстеру. Воплотить идею Пастера согласился один из его друзей врач Транше. Он считал, что в данном случае необходимо рискнуть. Риск оправдал надежды: мальчик не заболел. Вторым, кому прививки спасли жизнь, был четырнадцатилетний пастух Жан Батист Жюпиль, памятник которому стоит во дворце Пастеровского института в Париже. Через десять лет воскрешенные Пастером Ж. Мейстер и Ж. Б. Жюпиль были в первых рядах провожавших в последний путь великого ученого.
Обреченные на смерть люди, чтобы остаться в живых, любыми средствами и путями добирались до Парижа. Среди них были 16 крестьян из-под Смоленска и многие другие.
Первая в России и вторая в мире пастеровская станция была открыта в Одессе в 1886 г. Ее организаторами и руководителями были И. И. Мечников и Н. Ф. Гамалея. В настоящее время пастеровские станции, которые проводят прививки против бешенства, имеются во всех странах мира.
Луи Пастер сделал много открытий и тем самым способствовал прогрессу науки. Они представлены в лаконичной надписи у входа в его первую лабораторию в Париже.
«Здесь была лаборатория Пастера:
1857 г. – Брожения.
1860 г. – Самопроизвольное зарождение.
1865 г. – Болезни вина и пива.
1868 г. – Болезни шелковичных червей.
1881 г. – Зараза и вакцины.
1885 г. – Предохранение от бешенства».
В день своего семидесятилетия, обращаясь к ученым, приехавшим из разных стран мира, он сказал: «Вы доставляете мне самую большую радость, которую только может испытать человек, твердо верящий, что наука и мир восторжествуют над невежеством и войной, что народы найдут общий язык не для уничтожения, а для созидания, что будущее принадлежит тем, кто больше всех делает для страждущего человечества». Эти замечательные слова не потеряли своего значения и смысла и в наши дни.
Другим ученым, много сделавшим для усовершенствования методики работы с бактериальными культурами, был немецкий врач Роберт Кох (1843–1910). Он с сотрудниками впервые (1883) ввел в микробиологическую практику плотные питательные среды. На них можно выращивать и получать чистые культуры микроорганизмов, что
имеет большое значение при изучении неизвестных возбудителей болезней, Р. Кох обнаружил и изучил возбудителя туберкулеза человека и крупного рогатого скота (иногда возбудителя туберкулеза называют палочкой Коха).
Р. Кохом разработаны методы окрашивания микроорганизмов, микрофотографии, дезинфекции; введено в лабораторную практику заражение подопытных животных. Для подтверждения причины заразной болезни им была предложена методика, которая вошла в науку под названием триады Генле-Коха. Согласно этой методике необходимо:
1) установить, что микроб встречается только при определенной болезни;
2) выделить его в чистую культуру на искусственной питательной среде;
3) с помощью выделенного микроба вызвать у животного такое же заболевание.
Таким образом, Р. Кох заложил основы современной методики микробиологических исследований. В декабре 1905 г. Шведская академия наук присудила Р. Коху Нобелевскую премию.
Велика заслуга в развитии микробиологии нашего соотечественника И. И. Мечникова (1845–1916).
В результате многолетнего и упорного труда он создал фагоцитарную теорию иммунитета, в основу которого положена способность клеток организма противостоять инородным телам. К изучению иммунитета И. И. Мечников подошел с позиций сравнительной патологии. Клетки организма, окружая микробов или другие инородные тела, переваривают их и тем самым предохраняют организм от заражения. Такое явление получило название фагоцитоза и признано учеными всего мира.
Другой большой заслугой И. И. Мечникова является установление антагонизма между молочнокислыми и гнилостными микробами. На принципе антагонизма он обосновал теорию долголетия и предложил для продления человеческой жизни использовать простоквашу, которая впоследствии получила название Мечниковской. Гнилостные микробы, разлагая в кишечнике белки и другие вещества, образуют большое количество газов, таких, как аммиак, скатол, индол и др. Все они, являясь ядами, хотя и медленно, но отравляют организм, что ведет к сокращению человеческой жизни. Молочнокислые бактерии угнетают гнилостные, при этом количество продуктов распада уменьшается.
И. И. Мечников вторым среди русских ученых удостоен Нобелевской премии. Она ему присуждена (вместе с П. Эрлихом) в 1908 г. за работы по иммунитету (фагоцитарная теория). И. И. Мечников был членом академий наук почти всех стран мира. В 1888 г. он переехал в Париж, где в течение 28 лет работал в Пастеровском институте. В лаборатории Мечникова в Париже учились Д. К. Заболотный, Л. А. Тарасевич, И. Г. Савченко, Н. Я. Чистович, Ф. Я. Чистович, Г. Н. Габричевский, П. В. Циклинская и др. И. И. Мечников писал: «Моя лаборатория открыта для всех русских ученых, желающих работать и способных работать. Здесь они у себя дома».
Франция для И. И. Мечникова была второй родиной, а Париж – любимым городом, в котором он остался навсегда. Урна с его прахом замурована в стене библиотеки Пастеровского института.
Л. С. Ценковский (1822–1887) – один из основоположников отечественной микробиологии. По специальности ботаник, он занимался изучением простейших форм жизни. Родился в Варшаве. В 1844 г. он окончил Петербургский университет и через два года защитил диссертацию на ученую степень магистра бетаники. Л. С. Ценковский занимал должность профессора естественных наук в Ярославском лицее, а позже заведовал кафедрой ботаники в Петербургском университете. В 1856 г. он защитил докторскую диссертацию на тему: «О низших водорослях и инфузориях». В 1865 г. Л. С. Ценковский перешел на службу в Одессу, в 1872 г. переехал в Харьковский университет, где плодотворно работал в течение последних 15 лет. Самым ценным вкладом Л. С. Ценковского в науку этого периода является разработка методов прививок против сибирской язвы овец и других животных. В то время сибирская язва вызывала опустошительные эпизоотии. Поэтому предохранение животных от заболевания имело огромное значение для животноводства России. Вакцина против сибирской язвы, приготовленная и внедренная Л. С. Ценковским в России, через два года (1883) после подобных прививок Л. Пастера, была настолько совершенна, что в некоторой модификации почти на протяжении 80 лет служила средством предупреждения заболевания животных сибирской язвой. Л. С. Ценковский доказал бактериальную природу сахарного белка и разработал способы предупреждения его в сахарном производстве.
Д. И. Ивановский (1864–1920) – физиолог растений. Впервые открыл возбудителя мозаичной болезни табака (им оказался вирус). В 1892 г. в журнале «Сельское хозяйство и лесоводство» № 2 была опубликована его статья «О двух болезнях табака», а в 1903 г, в Киевском университете он защитил докторскую диссертацию на тему: «Мозаичная болезнь табака». Д. И. Ивановский доказал, что живое начало, проходящее через бактериальные фильтры, способно заражать здоровые листья табака и вызывать мозаичную болезнь. Это послужило толчком к открытию возбудителей ящура, оспы, невидимых в обычные световые микроскопы. Открыв вирус, Д. И. Ивановский стал основоположником новой ветви микробиологии — вирусологии.
С. Н. Виноградский (1856–1953) – основоположник почвенной микробиологии. Им проведены большие исследования по изучению серобактерий (1887), железобактерий (1888) и нитрифицирующих бактерий (1890). С. Н. Виноградский установил явление хемосинтеза. Это одно из крупных открытий в области физиологии микроорганизмов. Нитрифицирующие бактерии, которые окисляют аммиак до азотной кислоты, добывают углерод из углекислого газа воздуха, используя для этого процесса энергию, образуемую в результате химических реакций. Ученым предложены элективные (селективные, избирательные) среды, на которых можно культивировать определенные группы микроорганизмов и тем самым приближать их к естественным условиям обитания.
Н. Д. Иерусалимский (1900–1967), К А. Красильников (1896–1973), Е. Н. Мишутин (1901–1991) и др. внесли значительный вклад в развитие общей микробиологии. Ими написаны монографии и книги по разным разделам курса.
ТЕМА 3
МОРФОЛОГИЯ, СТРУКТУРНО-ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ПРОКАРИОТ.
Форма микробов. Микробы – это в основном одноклеточные бесхлорофилльные организмы прокариотического типа. По форме различают: шаровидные, палочковидные и извитые микробы (рис. 1). Между основными формами имеются и переходные (коккобактерии и др.).
Шаровидные (кокковые) микробы по форме напоминают шар, но бывают овальные, плоские, односторонне вогнутые или слегка вытянутые. Шаровидные формы образуются в результате деления клеток в одной, двух, трех взаимно перпендикулярных или разных плоскостях.
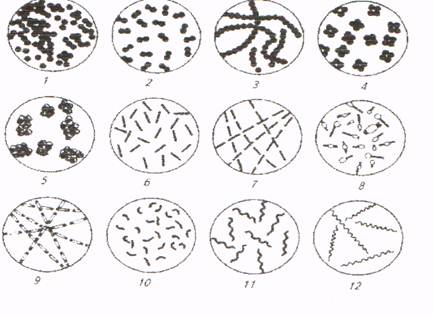
Рис.1. Основные формы микроорганизмов (схема):
шаровидные: 1 – стафилококки, 2 – диплококки, 3 – стрептококки, 4– тетракокки, 5 – сарцины; палочковидные: 6– бактерии, 7– стрептобактерии, 8– бациллы, 9 – стрептобациллы;
извитые: 10 – вибрионы, 11 – спириллы, 12 – спирохеты
такие формы получили название диплококков. Если деление происходит последовательно в одной плоскости и клетки соединены в виде цепочки, — это стрептококки. Деление кокка в двух взаимно перпендикулярных плоскостях ведет к образованию четырех клеток, или тетракокка. Пакетообразные кокки, или сарцины, — результат деления кокков в трех взаимно перпендикулярных плоскостях. Беспорядочное расположение клеток или образование скоплений, напоминающих гроздья винограда, происходит при делении кокков в разных плоскостях; такие формы называются стафилококками.
Палочковидные, ши цилиндрические, формы принято делить на бактерии и бациллы. Бактерии – палочковидные формы, не образующие спор (пишут Bad, например Bact. aceti). Бациллы – палочковидные формы, образующие споры (пишут Вас, например Вас. subtilis). Бактерии и бациллы бывают разными по форме и размерам. Концы палочек чаще закруглены, но могут быть срезаны под прямым углом (возбудитель сибирской язвы), иногда сужены. У мелких бактерий разница между длиной и шириной невелика; по внешнему виду они напоминают кокки, в связи с чем такие формы получили название коккобактерии (возбудитель бруцеллеза).
Спорообразующие микроорганизмы окрашиваются в основном по Граму положительно. Большинство из них имеют палочковидную форму и лишь Sporosarcina – шаровидную.
Среди палочковидных форм, образующих споры, различают бациллы и клостридии. Бациллы, за исключением Вас. anthracis, подвижны. Бациллы – аэробы, У бацилл споры не превышают толщины вегетативной клетки. Клостридии – анаэробы. Споры толще вегетативной клетки. Такие формы напоминают веретено, ракетку, лимон, барабанную палочку. Клостридии принимают участие во многих процессах в природе. Являются возбудителями анаэробных инфекций. Вызывают аммонификацию белковых веществ, мочевины. Разлагают фосфорорганические соединения. Фиксируют молекулярный азот и др.
Палочки, как и кокки, могут располагаться попарно или цепочкой. При соединении бактерий попарно образуются диплобактерии, при таком же соединении бацилл– диплобациллы. Соответственно образуются стрептобактерии и стрептобациллы, если клетки располагаются цепочкой. Тетрад и пакетов палочковидные формы не образуют, так как они делятся в одной плоскости, перпендикулярной продольной оси. Термин «бактерии» применяют для обозначения палочковидных форм, не образующих спор, и это правильно, в то время как многие авторы используют его как собирательное название разных микроорганизмов. Мы считаем, что вместо бактерии следует применять слово «микроорганизмы», или кратко «микробы». В учебнике «Основы микробиологии» В. Л. Омелянский отмечал, что понятие «микробиология» шире, чем «бактериология», так как оно охватывает собой не только бактерии, но и другие группы низших организмов: дрожжи, плесневые грибы, мелкие водоросли, которые называют микробами.
Извитые формы микробов определяют не только по длине и диаметру, но и по количеству завитков. Вибрионы напоминают по форме запятую. Спириллы– извитые формы, образующие до 3–5 завитков. Спирохеты – тонкие длинные извитые формы с множеством завитков. Они занимают промежуточное положение между бактериями и простейшими. Микобактерии – палочки с боковыми выростами (возбудители туберкулеза, паратуберкулеза). Коринебактерии напоминают микобактерии, но отличаются от них образующимися на концах утолщениями и включениями зерен в цитоплазме (дифтерийная палочка). Нитчатые бактерии — многоклеточные организмы, имеющие форму нити. Миксобактерии скользящие микробы, по форме напоминающие палочки
или веретено. Простекобактерии могут быть треугольной или иной формы. У некоторых: из них лучевая симметрия. Свое название такие организмы получили по наличию остроконечных выростов – простек. Размножаются они делением, или почкованием. Так, у треугольных форм на одной из вершин образуется почка, которая при достижении размеров материнской клетки отделяется. С помощью простек, расположенных на двух других вершинах, происходит улавливание пищи. Простекобактерии обычно неподвижны; подвижные формы образуют круговые движения. Спор не образуют, по Граму не окрашиваются. Растут на картофельной среде (агаре) при температуре 28 "С.
Размеры микробов. Микробы– микроскопические организмы. Их размеры определяются в микрометрах (мкм) (10~6 м по системе СИ). Диаметр шаровидных форм 0,7– 1,2 мкм; длина палочковидных 1,6—10 мкм, ширина 0,3—1 мкм. Вирусы — еще более мелкие существа. Их размеры определяются в нанометрах (I нм = 10~ м). Нитчатые формы микробов достигают длины в несколько десятков микрометров. Для того чтобы представить размеры этих существ, достаточно сказать, что в одной капле воды может вместиться несколько миллионов или миллиардов микробов.
Строение микробных клеток. Ввиду малых размеров строение микробов в течение долгого времени не было изучено. С появлением электронного микроскопа и ультрамикротома, при помощи которого можно делать очень тонкие срезы, представилась возможность глубже проникнуть в микромир. В результате было установлено, что микробные клетки имеют такое же сложное строение, как и клетки животных и растений (рис.2).

Микробная клетка окружена оболочкой. В ней различают капсулу, клеточную стенку и цитоплазматическую мембрану. Роль оболочки огромна: она придает клетке определенную форму, защищает ее от неблагоприятных воздействий, через нее осуществляется обмен веществ между клеткой и окружающей средой.
У некоторых микроорганизмов вокруг клеточной стенки образуется слизистый слой — капсула. У большинства микробов она состоит из полисахаридов, у отдельных содержит полипептиды и до 98 % воды. Капсула предохраняет клетку от высыхания, защищает ее от неблагоприятных воздействий макроорганизмов — фагоцитов, антител, обусловливает вирулентность. У возбудителя сибирской язвы капсула образуется в организме хозяина, а иногда также на средах с кровью, кровяной сывороткой животных.
Клеточная стенка – основная структурная единица оболочки микробной клетки. У «свободноживущих» микроорганизмов стенка представляет собой муреиновую сеть толщиной в 40 слоев. У грамотрицательных микробов муреиновая сеть однослойная (Г. Шлегель, 1987). Содержание пептидогликана (муреин, гликопептид, мукопептид) у грамположительных микробов составляет около 80 % массы сухого вещества стенки, у грамотрицатель-ных – 1–10 %. У грамположительных микробов клеточная стенка обычно толще и достигает 50 нм, у грамотрицательных – не превышает 5–15 нм. В клеточных стенках большинства грамположительных микробов содержится до 50 % (по массе) тейхоевых кислот, которые размещены по обе стороны пептидогликанового слоя. Их больше содержится на стороне, обращенной к цитоплазматической мембране, меньше– на наружной поверхности.
Микрофибриллы пептидогликана образуют каркас микробной клетки. При их переплетении формируются ячейки (поры), через которые осуществляется движение веществ внутрь клетки и из клетки во внешнюю среду. Разные химический состав и строение стенок клеток лежат в основе деления микробов на грамположительные и грамотрицательные организмы. При окрашивании по Граму основные красители (генциановый фиолетовый, кристаллический фиолетовый) в присутствии йода (раствор Люголя) с компонентами клетки образуют комплекс, который при действии на него этиловым спиртом удерживает краситель у грамположительных и обесцвечивает у грамотрицательных микробов. В результате грамположительные микробы окрашиваются в цвет основных красителей (фиолетовый, синий), а грамотрицательные — в красный (цвет дополнительного красителя — фуксина).
Цитоплазматическая мембрана, расположенная между клеточной стенкой и цитоплазмой, представляет собой относительно плотное образование, которое можно обнаружить при помощи электронного микроскопа. Мембрана состоит из фосфолипидного бислоя и белковых глобул. Отдельная молекула фосфолипидного слоя имеет форму головки с двумя отростками (вытянутые цепи жирных кислот). Молекулы расположены в два ряда, в которых гидрофильные головки направлены в противоположные стороны. Фосфолипидный слой образует структурную основу мембраны. Толщина его равна 4,5 нм (по Р. Капальди). По обе стороны бинарного фосфолипидного слоя расположены белковые глобулы (шарики) диаметром около 6 нм. Белки могут находиться на поверхности или погружаться в бислой фосфолипида. Одни из них пронизывают его насквозь, другие — на Уз своей величины (рис. 3). Белковые глобулы формируют каналы, которые выполняют функцию насосов
— с помощью АТФ осуществляют движение веществ и ионов (натрия, калия, кальция и др.).
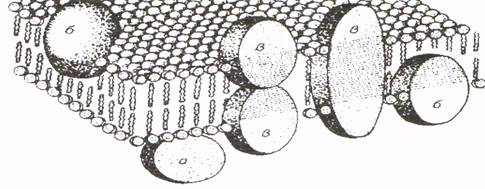
Рис. 3. Схема расположения слоев цитоплазматической мембраны
(по P.Капальди):
Глобулы белка: а – находятся на поверхности; б – погружены в фосфолипидный слой на небольшую глубину; в – пронизывают фосфолипидный слой поодиночке или парами.
Цитоплазма — содержимое клетки, за исключением ядра. Представляет собой коллоидную систему неодинаковой консистенции: чем ближе к поверхности, тем она плотнее. В цитоплазме содержатся рибосомы, мезосомы, включения (липиды, углеводы, волютин, сера, железо и другие соединения).
Рибосомы — рибонуклеопротеидные частицы цитоплазмы,, количество которых в одной клетке достигает нескольких тысяч. Сорма округлая или овальная, размер 20—25 нм. Располагаются одиночно или группами по 10—20 рибосом (полирибосома). Представлены двумя субъединицами: большой и малой. Имеют разный состав у прокариот и эвкариот. У прокариот состоят (по массе) на 2/3 из РНК и на 1/3 из белков. У звкариот они содержат примерно одинаковое количество РНК и белков. Коэффициент седиментации (осаждение) рибосом у прокариот 70S,
у эукариот — 80S. Рибосомы — место биосинтеза белка.
Мезосомы, или митохондрии, образуются в результате инвагинации (впячивания) и последующего ущемления цитоплазматнчеекой мембраны. Они являются энергетическими центрами клетки. С помощью ферментов в них происходит окисление жирных кислот, окислительное фосфорилирование, т. е. то, что называют процессами дыхания.
Гликоген — полисахариды, резервные вещества микробной клетки. Много гранулезы содержат маслянокислые бациллы. При действии на них раствором Люголя, в котором содержится йод, окрашиваются в синий цвет. Сера содержится в виде капель в клетках серобактерий. В такой же форме в клетках железобактерий находится железо.
Нуклеоид (генофор) — ядерное вещество прокариотической клетки, в отличие от ядер эукариот не имеет ядерной мембраны. Оно состоит из нуклеопротеидов, главным образом ДНК, принимает участие в передаче наследственных признаков и жизни клетки. Без нуклеоида клетка нежизнеспособна.
Споры (эндоспоры) образуются в конце экспоненциальной фазы роста, когда истощается питательная среда и накапливаются продукты жизнедеятельности. В это время внутри большинства вегетативных клеток появляются округлые образования, сильно преломляющие свет, которые отличаются от материнской клетки структурой, химическим составом и физиологическими свойствами. Эндоспоры образуют грамположительные анаэробные бациллы, грамположительные облигатно-анаэробные клостридии и грамположительные кокки рода Sporosarcina. У бацилл споры выполняют защитную функцию, у дрожжей и плесневых грибов служат для размножения. Форма спор округлая и овальная. Округлые споры чаще располагаются терминально (на концах клетки). Такие формы напоминают барабанные палочки (возбудитель столбняка). Овальные споры обычно располагаются в центре или ближе к одному из концов клетки и придают ей форму веретена — это клостридии (возбудитель эмфизематозного карбункула).
Жгутики — органы передвижения у микробов {рис. 6, а). Впервые обнаружены в 1838 г. Эренбергом и описаны в 1897 г. В. Мигулой. Они представляют собой тонкие спиральные нити, превышающие по длине размеры клетки. Микробы, имеющие прямую форму жгутиков, неподвижны. Диаметр жгутиков клетки выходит за пределы видимости в световом микроскопе, толщина их достигает 13 нм и более. Жгутик представляет собой цилиндр длиной до 20 мкм. В нем различают филамент (тело жгутика), крюк и базальное тело. Жгутик соединяется с клеткой базальным телом, которое имеет сложное строение. У основания жгутика находятся кольца. Нижним кольцом базальное тело прикрепляется к цитоплазматической мембране (рис. 6,). С помощью колец осуществляются вращательные движения. Установлено, что микробная клетка передвигается не волнообразными колебаниями жгутика, а его вращением (Берг, Андерсон, 1973), причем движение может быть как по ходу часовой стрелки, так и против хода часовой стрелки.
При множестве жгутиков они бывают собраны на конце клетки в один пучок и вытянуты вдоль продольной оси тела. Каждый из жгутиков пучка совершает синхронное и независимое вращательное движение. Вращается нижнее кольцо, находящееся в липидном слое клеточной мембраны, а вместе с ним и филамент. Такая система напоминает винт самолета, при вращении которого происходит движение микробной клетки.
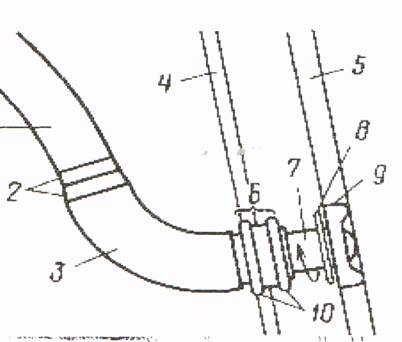
Рис. 7, Увеличенный жгутик бактерии Salmonella (а) и схема его строения (б) (по С. J. Jones, S. I. Апавд):
Сарцины — результат деления кокков в трех взаимно перпендикулярных плоскостях. (Сарцины) или колонии сферической формы. Спорообразующие микроорганизмы окрашиваются в основном по Граму положительно. Большинство из них имеют палочковидную форму и лишь Sporosarcina — шаровидную. Если же деление происходит в нескольких плоскостях неравномерно, образуются клеточные скопления неправильной формы. Прокариоты, имеющие форму цилиндра (палочковидные), сильно различаются по величине отношения длины клетки к ее поперечнику. Прокариоты спиралевидной формы отличаются разным количеством витков: у спирилл – от одного до нескольких витков, вибрионы выглядят наподобие изогнутых палочек, так что их можно рассматривать как неполный виток спирали. Многообразие и разнообразие форм: кольцо (замкнутое или разомкнутое в зависимости от стадии роста); образование простек – выростов от 1 до 8 (у м/о, размножающихся почкованием); в виде червей или 6-тиугольных звезд; в виде ветвящихся нитей. Выделяются и виды, имеющие морфологическую изменчивость в зависимости от условий: в виде палочек, кокков или слабого ветвления. У многоклеточных – это скопления различной конфигурации, чаще нити. Своеобразие бактериальным клеткам придают жгутики, имеющие различное расположение на клеточной поверхности, а также выделения внеклеточных веществ разной химической природы. Структура, химический состав и функции компонентов прокариотной клетки.. Принципиальные особенности ультраструктурной и химической организации: структуры, расположенные снаружи от ЦПМ – клеточная стенка, капсула, жгутики, ворсинки, слизистый чехол называют поверхностными структурами. Термином «клеточная оболочка» часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ – это капсула, клеточная стенка, слизистый чехол. Клеточная стенка – важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток не содержащих этих слоев клеточной оболочки). Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Клеточная стенка является определенным барьером для проникновения избытка воды в клетку как механическое образование, а содержание в клетке солей, создает осмотическое давление внутри клетки, которое и сохраняется благодаря наличию клеточной стенки и разности в осмотическом давлении в клетке и окружающей среде. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. Этот признак имеет резкие различия у прокариотной и эукариотной клеток. В зависимости от от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на 2-ве большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем иодом, образуется окрашенный комплекс. Если затем окрашенные препараты обработать спиртом, то у одних видов бактерий окраска сохраняется, а у других окрашенный комплекс разрушается и вымывается из клетки. Они обесцвечиваются. Этот способ был впервые предложен в 1884 г. датским ученым Х. Грамом, занимавшимся окраской тканей и позднее был использован для бактерий. И по отношению к этому способу окраски бактерии делятся на грамположительные и грамотрицательные виды. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки. Клеточные стенки грамположительных и грамотрицательных эубактерий резко отличаются как по химическому составу, так и по ультраструктуре. Отметим, что в состав клеточной стенки эубактерий входят 7 различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных эубактерий он составляет основную массу вещества клеточной стенки – от 40 до 90 %, у грамотрицательных его значительно меньше от 1 до 10 %, при том, что на долю клеточной стенки приходится от 5 до 50 % сухого вещества клетки. Различия в структуре клеточной стенки грамположительных и грамотрицательных эубактерий хорошо видны в электронном микроскопе. У первых – клеточная стенка выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий многослойная клеточная стенка, внутренний электронно-плотный слой которой состоит из пептидогликана и толщина его составляет 2 – 3 нм. Снаружи к нему прилегает, как правило, волнистый слой (8 – 10 нм), имеющий характерное строение: 2-ве электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому 3-хконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны. В состав пептидогликана входят углеводы N-ацетилглюкозамин и N-ацетилмурамовая кислота, остаток молочной кислоты, аминокислоты: L-аланин и D-глутаминовая и содержащая аминогруппу мезо-диаминопимелиновая карбоновая кислота. Отличительным признаком является и строение жгутика, которые располагаются на клеточной поверхности многих прокариот и являются структурами, определяющими способность клетки к движению в жидкой среде. Он состоит из 3-х частей: фибриллы – длинной спиральной нити, у поверхности клеточной стенки переходящей в утолщенную изогнутую структуру – крюк. С его помощью нить прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку. Базальное тело – это система из 2 или 4 колец нанизанных на стержень, являющийся продолжением крюка. 2-ва внутренних кольца (М и S) – обязательные составные части базального тела, в то время как наружные кольца (Р и L) отсутствуют у грамположительных эубактерий и следовательно не необходимы для движения. Последние кольца, имеющиеся только у грамотрицательных эубактерий, локализованы соответственно в пептидогликановом слое и в наружной мембране. Первое кольцо М локализовано в ЦПМ, второе в периплазматическом пространстве грамотрицательных или пептидогликановом мешке грамположительных эубактерий.
Клетка прокариот обладает рядом принципиальных особенностей. касающихся ее как ультраструктурных, так и химической организации. Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки) называют обычно поверхностными структурами. Клеточная оболочка – это все слои, распологающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). Клеточная стенка – важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с внешней средой (у клеток, не содержащих этих слоев клеточной оболочки). Функции клеточной стенки: механический барьер между протопластом и внешней средой; определяет форму клетки; в связи с наличием осмотического давления внутри клетки, клеточная стенка механически защищает клетку от проникновения в нее воды. Для понимания структуры клеточной стенки бактерий важно, прежде всего, указать на сходство ее опорного «скелета» с полимерами β-D-глюкозы – целлюлозой и хитином. Целлюлоза – главный компонент клеточных стенок у высших растений, водорослей, оомицетов. У бактерий целлюлоза в качестве материала клеточной стенки не встречается, однако она соединяет клетки Sаrсinа vеntriсuli в большие пакеты и придает пленке «уксусного гриба» (Мусоdеrmа асеti) прочную кожистую консистенцию, в результате выделений целлюлозы Асеtоbасtеr асеti subsр. хуlinum в виде тонких фибрилл в среду. Хитин – материал наружного скелета членистоногих и некоторых других животных. Он является важнейшим компонентом клеточных стенок большинства групп грибов (базидиомицетов, аскомицетов, зигомицетов). Структурными элементами хитина являются остатки N-ацетилглюкозамина, соединенные между собой подобно глюкозным остаткам в целлюлозе, т. е. 1,4-гликозидными связями. Однако в бактериальной стенке содержатся структуры и вещества, которых нет у животных и растений. Опорный скелет бактериальной стенки состоит в значительной степени из однородного полимера – пептидогликана муреина. Эта макромолекула – гетерополимер, построенный из цепочек, в которых чередуются остатки N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой β-1,4-гликозидными связями, они не входят в состав растительных и животных клеточных стенок. Такие неразветветвленные гетерополимерные цепи образуют основу муреина. Остатки мурамовой кислоты через лактильные группы соединены пептидной связью с аминокислотами – D-формами аланина и глутаминовой кислоты, а также не встречающаяся в составе белков мезо -диаминопимелиновая кислота. Отличительные особенности клеточных стенок грам-положительных и грам-отрицательных бактерий. Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений – тейхоевые кислоты, полимеры, построенные на основе рибита (5-и атомного спирта) или глицерина (3-х атомного спирта), остатки, которых соединены между собой фосфодиэфирными связями. Тейхоевые кислоты ковалентно могут быть связаны с N-ацетилмурамовой кислотой. Поскольку это длинные линейные молекулы, они могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки. В этом случае, вероятно, они являются основными антигенами грамположительных эубактерий. Для клеточной стенки также характерно наличие небольших количеств полисахаридов, белков и липидов. К N-ацетилмурамовой кислоте присоединен обычно короткий пептидный хвост, состоящий из небольшого числа (обычно 4 – 5) аминокислот, при этом две особенности пептидного хвоста заслуживают внимания. Первая особенность – наличие аминокислот в D-форме (неприродная конфигурация) и вторая – высоое содержание аминокислот с двумя аминогруппами. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы – в формировании пептидных связей между гетерополимерными цепчками. Нетрудно себе представить, что этим способом можно «сшить» между собой множество гетерополимерных цепей. Частота «сшивок» различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другим химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии. Это имеет принципиальное значение для пространственной организации пептидогликана. Пептидогликан, окружающий протопласт грамположительных эубактерий, – это по существу одна гигантская молекула, «сшитая» с помощью гликозидных и пептидных связей. Именно последние обеспечивают ей 3-хмерную пространственную организацию. Участвующие в образовании клеточных структур аминокислоты варьируют от вида к виду. Видоспецифическое строение опорного каркаса представляет собой хороший таксономический признак. У грам-отрицательных эубактерий строение клеточной стенки намного сложнее, чем у грам-положительных. В ее состав входит гораздо большее число макромолекул разного химического типа. Муреиновая сеть однослойная или 2-х слойная, составляет менее 10% сухой клеточной стенки и характеризуется весьма редкими поперечными связями между гетерополимрными цепями. Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Муреин содержит только мезо-диаминопимелиновую кислоту и не содержит лизина; межпептидные мостики отсутствуют, тейховые кислоты не обнаружены. Снаружи от пептидогликана располагается дополнительный слой клеточной стенки – наружная мембрана, состоящая из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида, они как бы наклеены снаружи на муреиновый каркас. Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80% всех наружной мембраны. Одна из функций этих белков – формирование в мембране гидрофильных пор диаметром примерно 1нм, через которые осуществляется неспецифическая диффузия молекул сахаров, аминокислот, небольших олигосахаридов и пептидов. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами.. Минорные белки наружной мембраны представлены гораздо большим числом видов, основной функцией которых является транспорт и рецепторная, например, белки, ответственные за специфический транспорт в клетку железосодержащих соединений.Таким образом, основные компоненты клеточной стенки эубактерий – три типа макромолекул: пептидогликаны, тейхоевые кислоты, полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией. Наличие в клеточных стенках пептидогликанового слоя – характерная особенность всех прокариот. Исключение составляют архебактерии, имеющие клеточную стенку, состоящую только из белка (экстремально галофильные, метанобразующие, ацидотермофильные), они не окрашиваются по Граму. Остальные типы архебактерий окрашиваются положительно по Граму. Именно, благодаря пептидогликану клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействия окружающей среды, обеспечивает поддержание ее внешней формы и дает возможность клетке существовать в гипотонических растворах. Организация клеточной стенки грам-отрицательных и грам-положительных бактерий имеет отличия по строению опорного муреинового каркаса и по содержанию других веществ. Так у грамотрицательных эубактерий клеточная стенка имеет отличия в структурной дифференцировке за счет формирования дополнительного слоя в виде наружной мембраны. Что значительно расширило круг функций клеточной стенки и, в первую очередь, за счет проблем проницаемости и транспорта веществ в клетку. Наружная мембрана имеет специфические и неспецифические каналы (поры) для пассивного транспорта веществ и ионов, необходимых клетке, т. е. осуществляет функции молекулярного «сита». Кроме того, наружная мембрана препятствует проникновению вредных веществ в клетку, чем можно объяснить большую устойчивость грамотрицательных бактерий (по сравнению с грамположительными) к ядам, ферментам, антибиотикам. Наличие дополнительной мембраны в составе клеточной стенки фактически обеспечило появление обособленной полости (периплазматического пространства), отграниченного от цитоплазмы и внешней среды специфическими мембранами и несущего важную физиологическую нагрузку, связанную с наличием в нем белков 2-х видов (транспортных и гидролитических ферментов). Грамположительные бактерии выделяют гидролитические ферменты во внешнюю среду, у грамотрицательных они локализованы в преиплазматическом пространстве. Разнообразные функции выполняют макромолекулы, локализованные частично или полностью на внешней стороне клеточной стенки, контактирующей с окружающей внешней средой; это специфические рецепторы для фагов и колицинов; антигены (липополисахарид грамотрицательных эубактерий, тейхоевые кислоты грамположительных; макромолекулы, обеспечивающие межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов. По компонентам и структуре клеточной стенки и биохимическим механизмам ее синтеза бактерии коренным образом отличаются от растений и животных. Лекарственные препараты, специфически воздействующие на клеточные стенки и на процесс их синтеза, должны быть безвредными для высших организмов. Действие лекарственных препаратов направлено, в первую очередь, на разрушение – лизис, клеточной стенки бактерий (пример, действие лизоцима, открытое А. Флемингом). Этим действием пользуются и микробиологи в практике получения протопластов и сферопластов. У первых клеточная стенка полностью лизируется и, следовательно, отсутствует; у вторых лизирована и отсутствует частично. В этих случаях также используют лизоцим, фермент из группы гликозидаз, который содержится в яичном белке, слезной жидкости и выделяется некоторыми бактериями. Полученные с помощью лизоцима сферопласты (из грамотрицательных эубактерий) или протопласты (из грамположительных) имеют сферическую форму и очень чувствительны к осмотическому давлению. Для их существования это давление должно быть сбалансировано с осмотическим давлением внутри клетки, при этом в них происходят процессы жизнедеятельности, но утрачивается способность к размножению. Однако, прокариоты, не имеющие клеточной стенки, обнаружены и в природе. Это группа микоплазм, сапрофитов и внутриклеточных паразитов растений, животных и человека. Формы, сходные с микоплазмами, были получены опытным путем с помощью вышеуказанных ферментов и антибиотиков, они носят название L-форм и образуются из нормальных бактериальных клеток в результате несбалансированного роста в длину и толщину. В благоприятных условиях они обладают метаболической активностью и способностью к размножению. Антибиотик пенициллин действует главным образом на грамположительные бактерии (стафилококки и пневмококки), а также на некоторые грамотрицательные клетки (гонококки, менингококки, энтеробактерии), причем только на растущие клетки, покоящиеся клетки не затрагиваются. Снаружи клеточная стенка прокариот часто бывает окружена оводненным веществом – капсулами и слизью. Жизненно важного значения для бактерий они не имеют, но некоторые бактерии в связи с этим имеют резистентность (от англ. rеsistоr – сопротивляюсь) к фагоцитозу, что повышает их вирулентность для животных. Капсула состоит в основном из полисахаридов (экзополисахаридов), относительно прочно связанных с клеточной стенкой и определяется с помощью окрашивания тушью, нигрозином, конго красным, в виде светлого ореола при микроскопировании окрашенных клеток. Капсулы с толщиной менее 0,2 мкм – микрокапсулы могут быть обнаружены только с помощью электронного микроскопа, с толщиной более 0,2 мкм – макрокапсулы, обнаруживаются световым микроскопом. Слизи – это выделения вещества капсул в окружающую среду, они не связаны с клеточной стенкой. Например, бактерия Lеuсоnоstос mеsеntеrоidеs – гетероферментативная молочнокислая бактерия, превращающая раствор, содержащий тростниковый сахар, в декстрановый студень, за что ее на сахарных заводах называют «бактерией лягушачьей икры». Декстран – это полисахарид, состоящий из остатков α-D-глюкозы, соединенных в положении 1,6 (1,6-α–глюкан), входит в состав кровезаменителей. Используется для повышения вязкости растворов и служит основой декстранового геля (сефадекса). Стрептококки, вызывающие кариес (разрушение зубов), в том числе Strерtососсus mutαns и S. sαlivαrius, выделяют другую гексозилтрансферазу, превращающую сахарозу в полифруктузы (леваны). Эти полисахариды откладываются на поверхности зубов и служат матриксом, в котором накапливаются кислые продукты брожения, вызываемого стрептококками, главным образом молочная кислота. В отличие от капсул чехлы имеют тонкую структуру и нередко в них обнаруживают много слоев с разным строением. Между указанными структурами у прокариот обнаружено много переходных форм, так что иногда нельзя четко отграничивать капсулу от слизистых клеточных выделений или капсулу от чехла. Наличие капсулы зависит от штамма микроорганизма и условий его культивирования, и образование капсулы может теряться в результате мутаций, таким образом капсулу нельзя рассматривать как обязательный структурный компонент клетки Расположение жгутиков: у палочковидных бактерий жгутики могут прикрепляться полярно (у полюсов или в полярной области клетки), если вдоль боковой поверхности – латерально. В зависимости от числа жгутиков и их локализации на поверхности клетки различают монополярные монотрихи, например, одним, но достаточно толстым жгутиком снабжены монотрихи – Vibriо mеtsсhnikоvii. Политрихи – бактерии с монополярным пучком жгутиков или биполярным жгутикованием в виде 2-х пучков жгутиков на каждом полюсе по пучку жгутиков (от 2 до 50). Лофотрихи – имеют много жгутиков на одном конце клетки, монополярно-политрихальное расположение жгутиков (Рsеudоmоnαs), биполярно-политрихальное называют амфитрихальным (Sрirillum) – амфитрихи. Перитрихи имеют жгутики по всей поверхности клетки или по бокам до 1000 на клетку. Толщина жгутика – 10 – 20 нм, длина 3 – 15 мкм. Вращение жгутика также как и его строение в виде спирали, закрученной против часовой стрелки, происходит против часовой стрелки. Структура мембран: мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные «головы» молекул обращены наружу, а гидрофобные «хвосты» погружены в толщу мембран. При «биологических температурах» мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. «Жидкая» структура мембран обеспечивает определенную свободу молекул белков, что является необходимым для осуществления процессов транспорта электронов и веществ через мембрану. Это же свойство обуславливает высокую эластичность мембран: они легко сливаются друг с другом, растягиваются и сжимаются. У мембранных белков, в отличие от липидов, нет единого способа структурной организации. Однако в зависимости от расположения в мембране и характера связи с липидным слоем мембранные белки условно можно разделить на 3-и группы: интегральные, периферические и поверхностные. Современная модель строения мембран учитывает большинство данных известных об их строении и представляет собой липидную основу, в которую включены ассиметрично расположенные белковые молекулы: интегральные белки образуют скопления на поверхностях липидного би- или монослоя, полностью погружены в него, а иногда пронизывают мембрану насквозь; периферические белки частично погружены в гидрофобную область; а поверхностные находятся вне её. Интегральные белки прочно связаны с мембранными липидами, периферические связаны с липидами в основном электростатическими силами, поверхностные белки связаны с мембранными липидами исключительно электростатическими взаимодействиями. В модели подчеркнута асимметрия строения мембраны, основанная на различиях в химическом строении и расположении молекул белка. Функции ЦПМ прокариот: барьерная – с помощью специальных переносчиков – транслоказ – через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов; энергетическая – процессы дыхания и фотосинтеза являются основными источниками энергии у бактерий, в ЦПМ расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (Δµн+), используемой затем в клетках по различным каналам, в том числе и для образования химической энергии (АТФ ); участие в репликации и последующем разделении хромосомы прокариотной клетки; интегрирующая роль, обеспечивающая определение клетки как единого целого. Избирательное поступление в клетку и выход из неё различных веществ и ионов осуществляется с использованием разных механизмов мембранного транспорта. Выделяют 4-е системы транспорта: пассивную диффузию; облегченную диффузию; активный транспорт и перенос химически модифицированных молекул. Движущей силой пассивной диффузии служит градиент концентрации вещества по обе стороны мембраны. Молекулы воды, некоторых газов О2, Н2, N2 и углеводородов, концентрация которых в о внешней среде выше, чем в клетке, проходят через ЦПМ, повидимому, через поры и с помощью переносчиков (транслоказ, или пермеаз, веществ белковой природы), локализованных в мембране и характеризующихся высокой субстратной специфичностью. Они связываясь с субстратом, подвергаются конформационным изменениям и вследствие этого приобретают способность к перемещению субстарта с одной стороны ЦПМ на другую. Облегченная диффузия происходит с участием транслоказ и не требует энергетических затрат и происходит только по градиенту концентрации. Наиболее распространен активный транспорт, позволяющий «накачивать» в клетку молекулы и ионы против их концентрационных и электрических градиентов. При этом для движения против электрохимического градиента требуются затраты метаболической энергии. Транспорт такого рода должен быть сопряжен с реакциями, продуцирующими энергию в химической или электрохимической форме. В результате переноса в виде химической модификации молекул, например, молекула глюкозы фосфорилируется и за счет этого обогащаются энергией. Внутриплазматические, (отличительные от наружной), мембраны прокариот: фотосинтетические, мезосомальные и прочие. Фотосинтетические мембраны характерны для фотосинтезирующих эубактерий (пурпурных, цианобактерий) и в клетке представлены в виде трубочек, пузырьков (везикул, хроматофоров) или уплотненных замкнутых дисков (тилакоидов), образованных 2-мя тесно сближенными мембранными пластинами (ламеллами). Локальные впячивания ЦПМ – мезосомы характерны для гра

