2014-10-30
2014-10-30 1595
1595РНК построена из тех же азотистых оснований, что и ДНК, но вместо тимина в ее состав входит урацил. Кроме того, углевод нуклеотидов РНК представлен рибозой.
Как происходит соединение нуклеотидов между собой в полинуклеотидной цепи?
В полинуклеотидной цепи соседние нуклеотиды связаны между собой ковалентными связями, которые образуются между дезоксирибозой (в молекуле ДНК) или рибозой (в молекуле РНК) одного нуклеотида и остатком фосфорной кислоты другого нуклеотида.
Чем объясняется огромное разнообразие генов в составе молекулы ДНК?
Хотя ДНК содержит всего четыре типа разных нуклеотидов, благодаря различной последовательности их расположения в длинной цепочке достигается огромное разнообразие их сочетаний в молекуле.
.
| Признаки | ДНК | РНК |
| Местонахождение в клетке | ядро, митохондрии, пластиды | ядро, цитоплазма, рибосомы, митохондрии, пластиды |
| Роль в клетке | химическая основа хромосомного генетического материала (генов); матрица для синтеза ДНК; матрица для синтеза РНК; информация о структуре белка | иРНК передает код наследственной информации о первичной структуре белка; рРНК входит в состав рибосом; тРНК переносит аминокислоты к рибосомам; митохондриальная и пластидная ДНК входят в состав этих органоидов |
| Строение | двойная спираль: две комплементарные полинуклеотидные цепи | одинарная полинуклеотидная цепь |
| Мономеры | дезоксирибонуклеотиды | рибонуклеотиды |
24
В 1950 г. английский физик М.Уилкинс получил рентгенограмму кристаллических волокон ДНК. Она показала, что молекула ДНК имеет определенную структуру, расшифровка которой помогла бы понять механизм функционирования ДНК. Рентгенограммы, полученные не на кристаллических волокнах ДНК, а на менее упорядоченных агрегатах, которые образуются при более высокой влажности, позволили Розалинд Франклин, коллеге М. Уилкинса, увидеть четкий крестообразный рисунок - опознавательный знак двойной спирали (рис. 13). Стало известно также, что нуклеотиды расположены друг от друга на расстоянии 0,34 нм, а на один виток спирали их приходится 10 (рис. 14). Диаметр молекулы ДНК составляет около 2 нм. Из рентгеноструктурных данных, однако, было не ясно, каким образом цепи удерживаются вместе в молекулах ДНК.
Картина полностью прояснилась в 1953 г., когда американский биохимик Дж. Уотсон и английский физик Ф. Крик, исследуя структуру молекулы ДНК, пришли к выводу, что сахарофосфатный остов находится на периферии молекулы ДНК, а пуриновые и пиримидиновые основания - в середине. Причем последние ориентированы таким образом, что между основаниями из противоположных Цепей могут образоваться водородные связи. Из построенной ими модели выявилось, что какой-либо пурин в одной цепи всегда связан водородными связями с одним из пиримидинов в другой цепи. Такие пары имеют одинаковый размер по всей длине молекулы. Не менее важно то, что аденин может спариваться лишь с тимином, а гуанин только с с цитозином. При этом между аденином и тимином образуются две водородные связи, а между гуанином и цитозином - три (рис. 15)
25
Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена большая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичным признаком, для которого характерен относительно низкий уровень индивидуальной изменчивости[1].
Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят о бактериальных или вирусных хромосомах. Поэтому, по мнению Д. Е. Корякова и И. Ф. Жимулёва[2], более широким определением является определение хромосомы как структуры, которая содержит нуклеиновую кислоту и функция которой состоит в хранении, реализации и передаче наследственной информации. Хромосомы эукариот — это ДНК-содержащие структуры в ядре, митохондриях и пластидах. Хромосомыпрокариот — это ДНК-содержащие структуры в клетке без ядра. Хромосомы вирусов — это молекула ДНК или РНК в составе капсида.
Морфология метафазных хромосом[править | править исходный текст]
В ходе клеточного цикла облик хромосомы меняется. В интерфазе это очень нежные структуры, занимающие в ядре отдельные хромосомные территории, но не заметные как обособленные образования при визуальном наблюдении. В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять свою целостность и форму[5][6]. Именно хромосомы на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа. Митотические хромосомы можно увидеть у любого организма, клетки которого способны делиться митозом, исключение составляют дрожжи S.cerevisiae, чьи хромосомы слишком малы[7]. Обычно митотические хромосомы имеют размеры в несколько микрон. Например, самая большая хромосома человека хромосома 1 имеет длину около 7 — 8 мкм в метафазе и 10 мкм в профазе митоза[8].
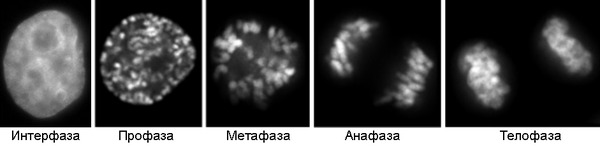

Клетки HeLa в интерфазе и на последовательных стадиях митоза
Метафазные хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. На стадии метафазы сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе[9]. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначается буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
· акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
· субметацентрические хромосомы с плечами неравной длины;
· метацентрические хромосомы, у которых центромера расположена посередине или почти посередине[10].
Эту классификацию хромосом на основе соотношения длин плеч предложил в 1912 году российский ботаник и цитолог С. Г. Навашин. Помимо вышеуказанных трёх типов С. Г. Навашин выделял ещё и телоцентрические хромосомы, то есть хромосомы только с одним плечом. Однако по современным представлениям истинно телоцентрических хромосом не бывает. Второе плечо, пусть даже очень короткое и невидимое в обычный микроскоп, всегда присутствует[11].
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомальные РНК. Небольшие хромосомные сегменты, отделяемые от основного тела хромосомы вторичными перетяжками, называются спутниками. Хромосомы, обладающие спутником, принято называть SAT-хромосомами (лат. SAT (Sine Acid Thymonucleinico) — без ДНК).