 2015-01-30
2015-01-30 2828
2828Дарвин считал, что видообразованию способствуют следующие условия: 1) большая плодовитость и широкое расселение вида в природе (чем больше рождается особей и чем в более разнообразных условиях они обитают, тем больше вариантов изменчивости и тем более разнообразным будет исходный материал для естественного отбора), 2) способность особей к насл
А щственной изменчивости, без которой невозможно возникновение отклонений для "испытания" отбором, 3) изоляция, в условиях которой обмена генов между популяциями не происходит и постепенно накапливаются изменения, отличающие изолированную популяцию от исходной.
В результате естественного отбора вид может измениться. От одной предковой формы могут также возникнуть- два и более видов. Процесс расхождения видов в разных экологических условиях Дарвин назвал дивергенцией. Примером ее могут служить вьюрки на Галапагосских островах (западнее Эквадора): одни питаются семенами, другие - кактусами, третьи - насекомыми. Каждая из этих форм отличается от другой величиной и формой клюва и могла возникнуть в результате дивергентной изменчивости и отбора. Еще более многообразны приспособления у плацентарных млекопитающих, среди которых встречаются наземные формы с быстрым бегом (собаки, олени), виды, ведущие древесный образ жизни (белка, обезьяна), животные, обитающие на земле и в воде (бобры, тюлени), живущие в воздушной среде (летучие мыши), водные животные (киты, дельфины) и виды с подземным образом жизни (кроты, землеройки). Все они происходят от единого примитивного предка - древесного насекомоядного млекопитающего.
Для естественного отбора имеет значение не выживание само по себе, а достижение организмом половозрелого возраста, успешное размножение и выживание потомства, которые и составляют основные критерии приспособленности, так как, размножаясь, половозрелая особь вносит материал в общий фонд наследственной основы вида. Таким образом, сущность дарвиновского естественного отбора составляют "перепроизводство" потомства и всеобщая случайная изменчивость, при этом шансы выжить и продолжить род имеют особи, отличающиеся от множества других какими-либо полезными свойствами. В борьбе за существование особи не обязательно должны погибнуть, они могут выжить, не оставив потомства, но их генотип исчезнет из общего генофонда. И, напротив, генотип преуспевающих "претендентов" становится массовым в данной популяции, давая основу для возникновения новых видов.
Рис. 90. Развитие различных типов приспособленности, берущих начало от сравнительно еще малоспециализированного организма: показано на представителях более высокоразвитых млекопитающих, произошедших от насекомоядных
ТАБЛИЦА (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
| Варианты видообразования | Способы, характерные для данного варианта |
| АЛЛОПАТРИЧЕСКОЕ (ГЕОГРАФИЧЕСКОЕ) | Новый вид возникает из одной или группы смежных популяций, расположенных на периферии ареала исходного вида |
| СИМПАТРИЧЕСКОЕ | Новый вид возникает внутри ареала исходного вида |
| ДИВЕРГЕНТНОЕ | Первоначальный единый вид разделяется на два или более в связи с возникновением межпопуляционных изоляционных барьеров и углублением различий под действием естественного отбора вплоть до генетической изоляции; увеличивается количество видов |
| ФИЛИТИЧЕСКОЕ | Новый вид возникает в результате постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп, что наблюдается при изменении условий во всем ареале |
| ГИБРИДОГЕННОЕ | Новый вид возникает путем гибридизации с последующим удвоением числа хромосом - аллополиплоидия |
Факторы (движущие силы) эволюции (Т.А. Козлова, В.С. Кучменко. Биология в таблицах. М.,2000)
| Фактор | Особенности проявления фактора |
| ЕСТЕСТВЕННЫЙ ОТБОР формы: движущий, стабилизирующий, дизруптивный | Избирательное (дифференциальное) воспроизведение генотипов (или генных комплексов), обеспечивающих выживание или гибель особей (см. табл. 34) |
| НАСЛЕДСТВЕННАЯ ИЗМЕНЧИВОСТЬ | Изменение генотипического состава популяций, генетическая гетерозиготность популяций обусловливается качественными перестройками генофонда; мутационный процесс рекомбинации - процессы, приводящие к возникновению новых геномов в результате перемешивания (рекомбинации) аллелей в популяциях; поток генов выражается в том, что в популяцию вносится новый источник изменчивости вместе с занесенной пыльцой или семенами других популяций, а также в результате скрещивания особей разных популяций; дрейф генов (генетико-автоматические процессы) происходит в популяциях, численность особей в которых оказывается ниже 500 - происходит гомозиготизация особей и затухание изменчивости |
| ИЗОЛЯЦИЯ пространственная (географическая); репродуктивная | Возникновение любых барьеров, препятствующих скрещиванию данного вида с другими видами; ее возникновение обусловлено наличием барьеров (водных, горных, особых участков суши и пр.) между ареалами разных видов; предотвращает межвидовые скрещивания, уменьшает успешность межвидовых скрещиваний (сезонная, механическая, этологическая, гибридологическая и др.) |
| ПОПУЛЯЦИОННЫЕ ВОЛНЫ | Периодические или апериодические колебания численности организмов в природных популяциях, изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны |
Вопрос
Макроэволюция — это процесс эволюционного преобразования и развития различных групп живых организмов на протяжении десятков и сотен миллионов лет. Иными словами, микроэволюция — это эволюционные преобразования живой природы на уровне выше видового (образование высших таксонов, новых органов и систем, вымирание отдельных групп и т. д.). В общем смысле макроэволюцией можно назвать развитие жизни на Земле в целом, включая и ее происхождение. Макроэволюционным событием считается также возникновение человека, по многим признакам отличающегося от других биологических видов. Между микро- и макроэволюцией нельзя провести резкую грань, потому что процесс микроэволюции, первично вызывающий изменение популяций (вплоть до видообразования), продолжается без какого-либо перерыва и на макроэволюционном уровне внутри вновь возникших форм.
Отсутствие принципиальных различий в протекании микро- и макроэволюционного процесса позволяет рассматривать их как две стороны единого эволюционного процесса, и применять для анализа процесса всей эволюции понятия, разработанные в теории микроэволюции, поскольку макроэволюционные явления (возникновение новых семейств, отрядов и других групп) охватывают десятки миллионов лет и исключают возможность их непосредственного экспериментального исследования.
Таким образом, можно представить следующие доказательства эволюционной теории.
Эмбрионологическое доказательство эволюции. Все многоклеточые животные проходят в ходе индивидуального развития стадии бластулы и гаструлы. С особой отчетливостью выступает сходство эмбриональных стадий в пределах отдельных видов и классов. Например, у всех наземных позвоночных, так же как и у рыб, обнаруживается закладка жаберных дуг, хотя эти образования не имеют функционального значения у взрослых организмов. Подобное сходство эмбриональных стадий обьясняется единством происхождения всех живых организмов.
Морфологическое доказательство эволюции. Существование форм, в которых сочетаются признаки нескольких характерных систематических единиц указывает на то, что в прежние геологические эпохи жили организмы, которые являются родоначальниками нескольких систематических групп. Связь между разными классами животных так же хорошо иллюстрирует общность их происхождения.
Эволюционное развитие может идти разными путями в зависимости от объекта и соотношения его со средой. Выделяют несколько форм исторического преобразования того или иного таксона.
Под филетической эволюцией понимают постепенные эволюционные перестройки определенной таксономической группы, которые ведут к возникновению новой труппы, отличной от исходной. Примером филетической эволюции может быть ряд предков лошади: фенокодус — эогиппус — миогиппус — парагиппус — плиогиппус — современная лошадь.
Дивергентная эволюция обусловливает распад одного таксона на несколько групп в процессе приспособления их к разным.условиям существования (рис. 100). Дивергентная эволюция осуществляется на разных уровнях. Она расчленяет вид на популяции, которые при возникновении изоляции могут превратиться в новые виды. Виды одного
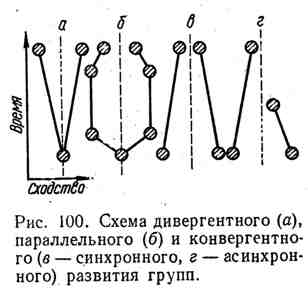
рода или роды одного семейства — результат дивергентной эволюции. Дивергенция любого масштаба увеличивает разнообразие форм жизни.
Дивергенция внутри вида — явление обратимое: две разошедшиеся популяции путем скрещивания могут объединиться. Макроэволюционные изменения необратимы. Формирование таксонов разного ранга в процессе дивергенции направляется групповым естественным отбором.
Параллелизм. Если ранее дивергировавшие группы попадают в сходные условия существования, у них независимо, параллельно могут формироваться сходные адаптивные признаки. Так, у глубоководных рыб параллельно и независимо возникли органы свечения. Независимое развитие сходных признаков у одновременно существующих родственных групп называют синхронным параллелизмом.
Таким образом происходили преобразования пятипалой конечности в однопалую у литоптерн Южной Америки и у лошадиных Северной Америки и Азии. При асинхронном параллелизме наблюдается независимое приобретение сходных черт близкими группами, которые жили в разное время. Много примеров такого асинхронного развития дает палеонтология. Показано, что саблезубость в семействе кошачьих возникала четырежды в двух независимых стволах.
Конвергенция, Неродственные группы, развиваясь в сходных условиях, могут приобретать сходные признаки. Конвергентное развитие можно наблюдать на всех уровнях: сходство форм тела у акул, ихтиозавров и дельфинов, общие черты строения тела у прыгающих млекопитающих, конвергентное внешнее подобие у многих форм сумчатых и плацентарных млекопитающих и т. д.
Вопрос

