 2015-01-21
2015-01-21 1069
1069Среди РНК различают: информационную, рибосомальную, транспортную, регуляторную, гетерогенно ядерную и др. На долю транспортной РНК (тРНК) приходится 10-15% всех РНК клетки, молекулярная масса не превышает 30 тыс. Д, ее нить состоит из 70-95 мононуклеотидов, причем в ее молекулу включено до 8-19% минорных оснований (дигидрацил, инозин, тиоурацил и т. д.). Необычность их структуры мешает нуклеазам разрушать тРНК, что делает их долгожителями. Оба конца нити тРНК у всех их представителей имеют одинаковое строение: в начале цепи расположен ГМФ, а заканчивается следующим тринуклеотидом: ЦЦА. Вторичная структура представлена в форме клеверного листа, образованной в виде шпилькообразных структур, за счет комплементарности (схема 3.2.3.1.1.). Сближенные концы цепи тРНК называют акцепторным участком, сюда крепится транспортируемая аминокислота, противоположная петля включает антикодон – триплет, с помощью которого узнается соответствующее (комплементарное) звено на иРНК. Исходя из того, что смысловых кодонов 61, следовательно, и разных тРНК в клетке не менее 60. Еще две петли служат для взаимодействия с рибосомой и аминоацил-тРНК-синтетазой. Цепи различных тРНК укладываются в компактные третичные структуры; за счет стекинг - взаимодействий петли сближаются, принимая форму, напоминающую латинскую букву «L». Митохондриальные тРНК несколько короче (до 65 нуклеотидов), петли также меньше, пространственная структура
не установлена.

Схема 3.2.3.1.1. Универсальная укладка полинуклеотидной цепи тРНК.
Функции т-РНК: а) акцепторная – способность ковалентно связываться с аминокислотой, превращаясь в аминоацил~тРНК; б) адапторная – способность узнавать кодом иРНК, соответствующий транспортируемой аминокислоте, и обеспечивать ее включение на законное место в растущей цепи белка. При этом образование пептидной связи обеспечивается за счет макроэргической связи аминоацил~т-РНК.
Большая часть цитоплазматических РНК (80-85%) составляют рибосомальные (рРНК), составляющие основу соответсвующих органелл, локализирующихся и в цитозоле, и в митохондриях. Выделяют несколько рРНК (5S, 16-18S, 23-29S) с различными ММ (от 40.000 до 700.000 Д), с неодинаковым числом мононуклеотидов (120-3000). Первичная структура включает минорные основания, хотя и в меньшем количестве, чем у тРНК.
Дальнейшее усложнение организации приводит к спирализации (с помощью водородных, координационных связей), возникновению шпилек с дефектами комплементарности (пиримидин - пиримидин). С помощью шпилек рРНК связываются со своими белками, обычно гистонами, образуя в целом рибосомы, предварительно плотно упаковавшись в пространстве (третичная структура), принимая разные формы (У или V). Функция рибосом – создать каркас, где бы происходило взаимодействие иРНК и тРНК и рост полипептидной цепи. Матричная (информационная) РНК (иРНК) резко отличается от других вариантов полирибонуклеотидов по структуре. На ее долю приходится от 2 до 6% от числа подобных соединений, молекулярная масса колеблется от 25000 до миллиона Д; может включать от сотни до 3000 мононуклеидов. В первичной структуре отсутствуют минорные основания, с одной стороны цепь заканчивается
 |
Схема 3.2.3.1.1. Схема первичной структуры и-РНК.
одинаковыми кирпичиками (50-400 моноаденилатов), подобный неспецифический хвост мешает разрушению иРНК нуклеазами. С другого конца имеет иРНК модифицированный ГМФ (сар-участок), с помощью которого крепится к рибосоме (схема 3.2.3.1.1.). В середине молекулы между инициирующим и терминирующим кодонами располагается собственно кодирующая область – представляющая чередование информативных и неинформативных участков. Вторичная и третичная структуры индивидуальны, но в итоге имеется несколько шпилек, куда крепятся факторы инициации и терминации, а молекула обязательно связывается со специфическим белком, образуя информосому, сближается с рибосомой, где служит матрицей в синтезе соответствующей полипептидной цепи.
Варианты ДНК
Дезоксирибонуклеиновые кислоты (ДНК) обычно локализуются в ядре и митохондриях, что и лежит в основе их деления.
Принципы строения их молекул однотипны. Как любой биополимер имеют несколько уровней организации: первичную – двойную полинуклеотидную нить, состоящую из дезоксирибонуклеозидфосфатов, связанных фосфодиэфирными связями; вторичную – полученную в результате спирализации; третичную – суперспиралированную структуру. Но так как они сильно отличаются по массе, то и в деталях их строения имеется много отличий.
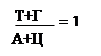
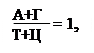
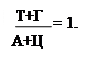 Митохондриальную ДНК иногда называют 25–ой хромосомой, ее значения не превышают 5% от общего количества ДНК; в каждой органелле содержится 5-10 ее копий. Это довольно маленький биополимер: его первичную структуру составляют 16569 пар мононуклеотидов. Облигатной чертой ДНК эукарит является спаренность нитей за счет комплементарности. Первым эту особенность заметил Chargaff в 1949 г. Он обратил внимание, что часть и пиримидиновых, и пуриновых оснований включает или кето- или аминогруппы, между которыми за счет наличия водорода в последней могут образовываться водородные связи. Его первый постулат: молярная доля пуринов равна молярной доле пиримидинов и это логично – две нити будут параллельны только тогда, если против крупной структуры (пурина) встанет маленькая (пиримидин). Если подобная ситуация создастся между двумя пуринами – им будет тесно, а пиримидинам – просторно, что не будет способствовать возникновению водородных связей. Во-вторых, для этого необходимо, чтобы против кетогруппы стояла аминогруппа, то есть их число должно быть равным
Митохондриальную ДНК иногда называют 25–ой хромосомой, ее значения не превышают 5% от общего количества ДНК; в каждой органелле содержится 5-10 ее копий. Это довольно маленький биополимер: его первичную структуру составляют 16569 пар мононуклеотидов. Облигатной чертой ДНК эукарит является спаренность нитей за счет комплементарности. Первым эту особенность заметил Chargaff в 1949 г. Он обратил внимание, что часть и пиримидиновых, и пуриновых оснований включает или кето- или аминогруппы, между которыми за счет наличия водорода в последней могут образовываться водородные связи. Его первый постулат: молярная доля пуринов равна молярной доле пиримидинов и это логично – две нити будут параллельны только тогда, если против крупной структуры (пурина) встанет маленькая (пиримидин). Если подобная ситуация создастся между двумя пуринами – им будет тесно, а пиримидинам – просторно, что не будет способствовать возникновению водородных связей. Во-вторых, для этого необходимо, чтобы против кетогруппы стояла аминогруппа, то есть их число должно быть равным
Исходя из выше сказанного, третьим положением и будет: А = Т, а Г = Ц (схема 323.2.1).
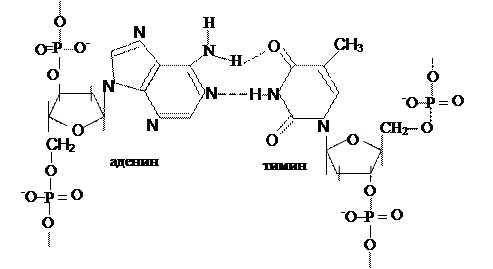
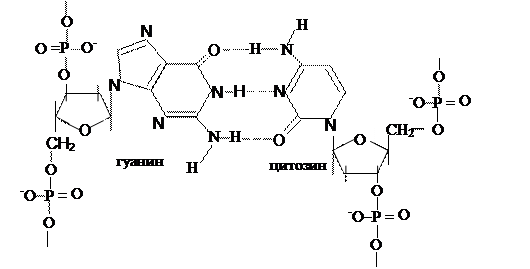
Схема 3.2.3.2.1. Водородные связи между основаниями в ДНК.
В целом, первичная структура ДНК представляет нить, образованную фосфодиэфирными связями между мононуклеотидами одной цепи и водородными между противоположными. (см. Приложение, схема 2). Кроме того, рядом лежащие гидрофобные ароматические основания за счет различных по плотности электронных облаков способны взаимодействовать друг с другом, создавая так называемые стекинг-взаимодействия, а фосфаты, являясь заряженными группами, вступают в ионные связи с различными катионами (металлов, основными белками, полиаминами).
Характеризуя митохондриальную ДНК следует заметить, что в первичной структуре отсутствуют интроны, в ее кодогенах (триплетах - комплементарных смысловым кодонам иРНК) нередки отклонения от генетического кода. Вторичная структура - хорошо спирализованное образование. Третичная - представляет кольцевой дуплекс, то есть концы сдвоенной нити сближены и соединены между собой. ДНК митохондрий содержит информацию не обо всех белках этих органоидов (см. «Патохимия наследственных болезней» С. 18).
Самая большая молекула, содержащая в клетках, это ядерная ДНК. Если ее вытянуть до первичной структуры, длина составит около 8 см (!). Этот уровень организации включает различные гены (транскриптоны, регуляционные и т.д.), палиндромы. Последние представляют фрагменты нитей, одна половина которой комплементарна второй, при их сближении друг с другом за счет возникновения водородных связей образуются шпильки, к которым крепятся различные белки (регуляторные, каталитические).
Первичная структура включает и минорные основания, о вероятной роли которых было написано выше.
Как замечено раньше, нить с одного конца фосфорилирована (5’ –конец), на другом расположена свободная 3’–ОН группа дезоксирибозы (3’ – конец). Когда две нити связаны одной ДНК, то они антипараллельны - каждый конец цепи имеет 3’ и 5’ концы.
Ядерная ДНК хорошо спирализована - это правозакрученная спираль, шаг которой равен 3,4 нм, диаметр составляет 2 нм, 1 виток включает 10 мононуклеотидов и полюс шпильки. Подобная конформация известна как В-форма и обычна для клетки. Но молекула ДНК может менять свою упаковку в зависимости от условий: быть более сплющенный (А-форма), зигзагообразный (Z-форма).
Важным свойством двойной спирали ДНК является ее способность изгибаться. Молекула ее в миллион раз длиннее, чем ядро или клетка и чтобы поместиться в них, она должна быть гибкой, что приводит к суперспирализации, т.е. третичная структура представляет дальнейшее закручивание двойной нити вокруг положительно заряженных белков - гистонов. Последние образуют октамерный (восемь, т.е 4 пары Н2а, Н2в, Н3, Н4 гистонов) протеиновый комплекс, называемый нуклеосомой, вокруг которого молекула ДНК совершает два оборота (схема 3.2.3.2.2).
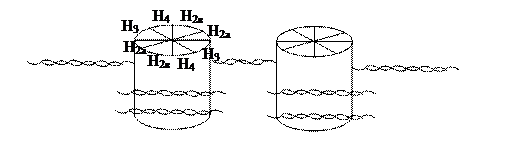
Схема 3.2.3.2. Модель упаковки ДНК
Участок ДНК, расположенный между рядом лежащими нуклеосомами и включающий 30-40 пар мононуклеотидов (в отличие от 200 пар, окружающих катушку), - называют линкерным. Считают, что он связан с Н1-гистонами. Подобная упаковка доводит до стократного уменьшения молекулы. В дальнейшем подобные нити образуют петли, что увеличивает компактность; за счет дополнительной спирализации и обеспечивается 10000-кратная конденсация ДНК.








