 2015-02-27
2015-02-27 31321
31321Автор: преподаватель, Д. Г. Кнорре, Преподаватель, Высшая школа. Тип материала: Учебник
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины. Последняя выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80–85% от всего азота мочи. Основным и, возможно, единственным местом синтеза мочевины является печень. Впервые Г. Кребс и К. Гензеляйт в 1932 г. вывели уравнения реакций синтеза мочевины, которые представлены в виде цикла, получившего в литературе название орнитинового цикла мочевинообразования Кребса.
Следует указать, что в биохимии это была первая циклическая система метаболизма, описание которой почти на 5 лет опеределило открытие Г. Кребсом другого метаболического процесса – цикла трикарбоновых кислот. Дальнейшие исследования в основном подтвердили циклический характер биосинтеза мочевины в печени. Благодаря исследованиям Г. Коена, С. Ратнер и сотр. были уточнены промежуточные этапы и ферментные системы, катализирующие образование мочевины.
Таким образом, весь цикл мочевинообразования может быть представлен следующим образом. На первом этапе синтезируется макроэргическое соединение карбамоилфосфат – метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза пиримидиновых нуклеотидов (соответственно ДНК и РНК) и аргинина (соответственно белка и мочевины):
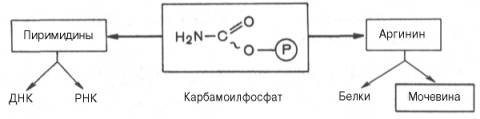
Мочевина – основной конечный продукт азотистого обмена, в составе которого из организма выводится избыток азота.
Орнитиновый цикл в печени выполняет две функции:
- превращение аминокислот в мочевину, которая экскретируется и предотвращает накопление токсичных продуктов, главным образом аммиака;
- синтез аргинина и пополнение его фонда в организме.
Рассмотрим цикл мочевины:
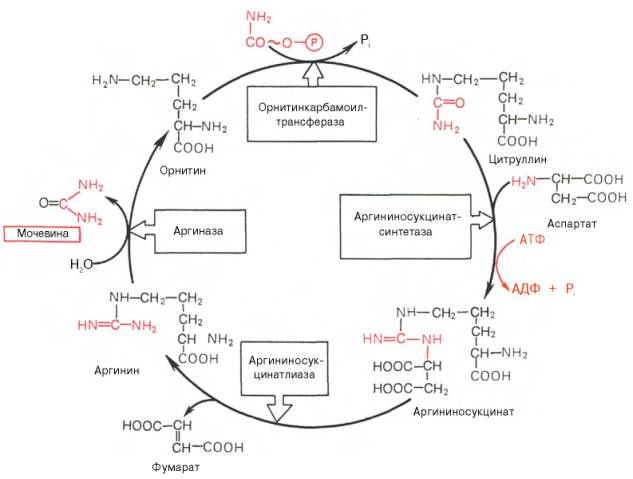
Рисунок 34 - Орнитиновый цикл синтеза мочевины в печени
Начинается он с образования карбамоилфосфата в митохондриях, где много АТФ.
1 Образование карбамоилфосфата. Ионы аммония, возникшие в результате окислительного дезаминирования глутаминовой кислоты, взаимодействуют с гидрокарбонат - анионом и АТФ при участии карбамоилфосфатсинтетазы, образуя карбамоилфосфат, содержащий макроэргическую связь:
О
//
NН4+ + НСО3- + 2АТФ → Н2N – С + 2АДФ + Н2РО4- + Н+
\
О ~ Ф
Карбамоилфосфат
2. Получение цитруллина. В матриксе митохондрий карбамоилфосфат конденсируется с аминокислотой орнитином, которая, являясь гомологом лизина, не входит в состав белков.
Реакция катализируется орнитинкарбамоилтрансферазой:
Н СОО- О Н СОО-
\ / // \ /
С + Н2N – С → СО + Н2РО4-
+ / \ + \ + / \ ||
Н3N (СН2)3NН3 О ~ Ф Н3N (СН2)3NН – С – NН2
Орнитин Карбамоилфосфат цитруллин
Образующийся цитруллин переходит в цитозоль клеток печени, где и происходят остальные реакции цикла мочевины.
3. Получение аргининосукцината. Замещение карбонильной группы цитруллина на аминогруппу аспартата с образованием гуанидиновой группировки аргининосукцината происходит при участии АТФ и катализируется аргининосукцинатсинтетазой:
Н СОО- Н СОО-
\ / \ /
С О + С + АТФ →
+ / \ || / \
Н3N (СН2)3NН – С – NН2 Н3N СН2СОО-
Цитруллин аспартат
Н СОО-
\ / +
С Н2N СН2СОО- + Н2Р2О72- + АМФ
+ / \ || |
Н3N (СН2)3NН – С – NН – СНСОО-
Аргининосукцинат
Реакция эндэргоническая, на протекание первой и третьей реакций цикла расходуется 4 молекулы АТФ.
4. Распад аргининосукцината. Под действием аргининосукцинатлиазы аргининосукцинат экзэргонически расщепляется с образованием аргинина и фумарата:
Н СОО- Н СОО- СОО- Н
\ / + \ / + \ /
С Н2N СН2СОО- → С Н2N + С = С
+ / \ || | + / \ || / \
Н3N (СН2)3NН – С – NН – СНСОО- Н3N (СН2)3NН – С – NН2 Н СОО-
Аргининосукцинат Аргинин Фумарат
5. Образование мочевины и регенерация орнитина. Гидролиз аргинина, катализируемый аргиназой, приводит к образованию мочевины и регенерации орнитина. Реакция экзэргонична.
Н СОО- Н СОО- NН2
\ / + + \ / /
С Н2N → С + О = С
+ / \ || + / \ + \
Н3N (СН2)3NН – С – NН2 Н3N (СН2)3NН3 NН2
Аргинин Орнитин Мочевина
Регенерированный орнитин может снова поступать в митохондрии и участвовать в новом обороте цикла мочевины. Образующуюся мочевину кровь переносит из печени в почки, где извлекается из крови и удаляется из организма с мочой.
В орнитиновом цикле расходуется 4 макроэргические связи трех молекул АТФ на каждый оборот цикла.
Однако процесс образования мочевины обеспечивает сам себя энергией:
- при регенерации аспартата из фумарата на стадии дегидрирования малата образуется НАДН, который может обеспечить синтез 3 макроэргических связей.
- при окислительном дезаминировании глутамата в разных органах тоже образуется НАДН, который может обеспечить синтез 3 макроэргических связей.
Из приведенных реакций видно, что токсичный аммиак превращается в безвредную мочевину. При этом один из атомов азота мочевины образуется из аммиака, другой – из аспартата.

