 2015-02-04
2015-02-04 986
986
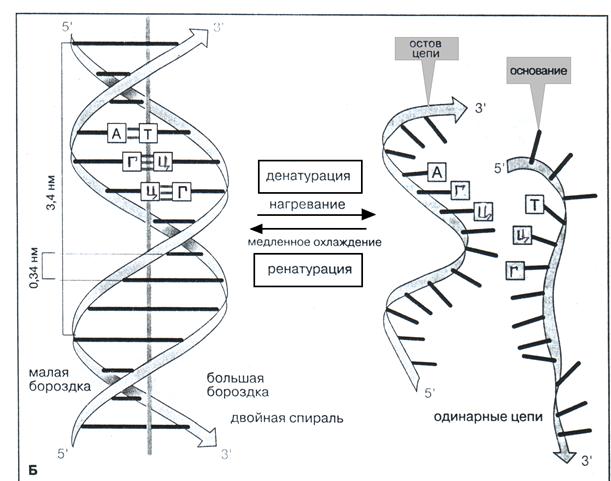
Рис. 3. Вторичная структура ДНК
Спаривания оснований А/Т и Г/Ц и структура в виде двойной спирали ДНК возможны только в том случае, если цепи ДНК располагаются антипараллельно, т.е. имеют противоположную направленность (полярность). Цепь, имеющую ориентацию фосфатных мостиков 3′ → 5′, т.е. полярность 5′р---...3′(ОН), называют правой и обозначают как «+» цепь. Цепь с противоположной ориентацией фосфатных мостиков 5′ → 3′, т.е. полярностью (ОН)3′----.....р5′ называют левой и обозначают как «–» цепь (рис.4).
Обратите внимание, что цепи ДНК отличаются друг от друга по составу азотистых оснований, а также по их последовательности, если каждую цепь читать в направлении от 5′р-конца к 3′(ОН)-концу. Кроме того, соблюдается равенство А и Т, Г и Ц (в каждом типе ДНК содержится примерно одинаковые количества А и Т, тоже самое относится к Г и Ц).
(+) 5′рА-А-Ц-Ц-Т- Г-А-Ц-Ц-А-Ц-Т- Г-А-Т-....3′(ОН) Фрагмент
(-) (ОН)3′ Т-Т- Г-Г-А-Ц-Т- Г- Г-Т- Г-А-Ц-Т-А-....р′5 ДНК
Принцип антипараллельности цепей ДНК имеет глубокий биологический смысл, так как в функциональном отношении две цепи не эквивалентны. Кодирующей цепью (матричной, смысловой) является (+) цепь, которая узнается транскрибирующим ферментом (РНК-полимеразой) и считывается в процессе транскрипции ДНК.
В зависимости от длины витка и расположения азотистых оснований по отношению к оси двойной спирали (угла спирали), различают более десяти разнообразных упорядоченных спиральных структур ДНК. Преобладающая в клетке конформация ДНК (так называемая В-ДНК) представлена на рис. 3Б. Длина витка (шаг спирали) составляет 3,4 нм, угол спирали - 90°. В стабилизации вторичной структуры ДНК наряду с водородными связями (Н-связи) большую роль играют гидрофобные взаимодействия между пространственно сближенными плоскостями азотистых оснований, направленные вдоль оси двойной спирали. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, из называют стэкинг-взаимодействиями (в В-форме ДНК расстояние между плоскостями азотистых оснований составляет 0,34 нм, т.к. в витке находится 10 нуклеотидных пар). Сильное стэкинг-взаимодействие всегда усиливает Н-связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, полярные фосфатные группы которого находятся на поверхности спирали. Внутренняя область двойной спирали, где расположены основания, неполярна. При ослаблении сил стэкинг- взаимодействия молекулы воды, проникая внутрь двойной спирали, конкурируют с полярными группами оснований за образование Н-связей, что в свою очередь, приводит к нарушению Н-связей между комплементарными парами оснований и уменьшению эффективности стэкинг-взаимодействия между азотистыми основаниями, инициирует дестабилизацию двойной спирали и способствует расхождению цепей (денатурации ДНК). Ферментативное локальное расхождение цепей ДНК имеет место в генетических процессах, связанных с воспроизведением и экспрессией наследственной информации. После окончания воздействия компонентов окружающего раствора цепи ДНК, благодаря спариванию оснований, вновь образуют двойную спираль в участке их расхождения (ренатурация ДНК). Все это свидетельствует о динамичности вторичной структуры ДНК. Процессы денатурации и ренатурации ДНК играют важную роль в генной инженерии.
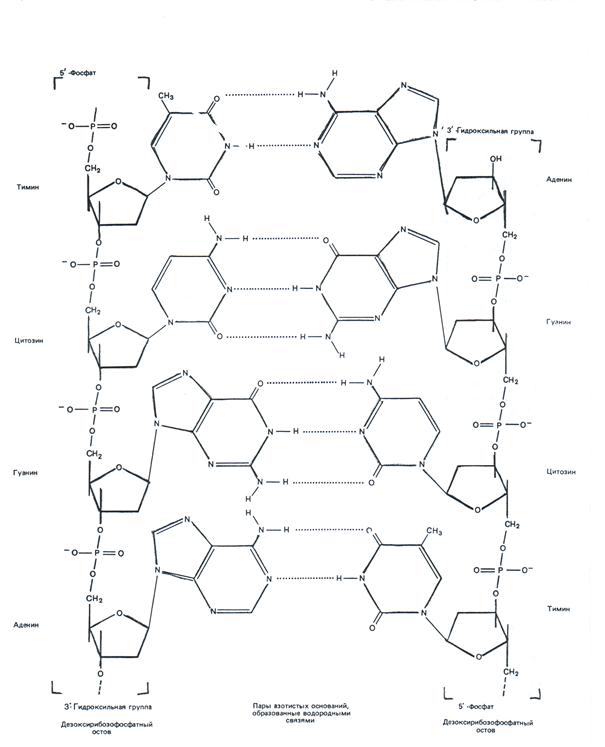
Рис. 4. Природа генетической информации (цепи ДНК комплементарны и антипараллельны)








