 2015-03-20
2015-03-20 7442
7442• Возбудимость - способность приходить в состояние возбуждения при действии раздражителей.
• Проводимость - способность проводить возбуждение.
• Сократимость - способность мышцы изменять свою длину или напряжение в ответ на действие раздражителя.
• Лабильность - лабильность мышцы равна 200-300 Гц.
При непосредственном раздражении мышцы (прямое раздражение) или опосредовано через иннервирующий ее двигательный нерв (непрямое раздражение) одиночным стимулом возникает одиночное мышечное сокращение, в котором выделяют три фазы:
• латентный период - время от начала действия раздражителя до начала ответной реакции;
• фаза сокращения (фаза укорочения);
• фаза расслабления.
В естественных условиях к скелетной мышце из ЦНС поступают не одиночные импульсы, а серия импульсов, следующих друг за другом с определенными интервалами, на которую мышца отвечает длительным сокращением. Такое длительное сокращение мышцы, возникающее в ответ на ритмическое раздражение получило название тетанического сокращения или тетануса. Различают два вида тетануса: зубчатый и гладкий.
Если каждый последующий импульс возбуждения поступает к мышце в тот период, когда она находится в фазе укорочения, то возникает гладкий тетанус, а если в фазу расслабления - зубчатый тетанус (рис. 5).
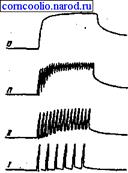
Рис. 5. Различные виды тетануса при повышении частоты раздражения. I - одиночные сокращения; II-III - зубчатый тетанус; VI - гладкий (сплошной) тетанус
Амплитуда тетанического сокращения превышает амплитуду одиночного мышечного сокращения. Исходя из этого Гельмгольц объяснил процесс тетанического сокращения простой суперпозицией, т. е. простой суммацией амплитуды одного мышечного сокращения с амплитудой другого. Однако в дальнейшем было показано, что при тетанусе имеет место не простое сложение двух механических эффектов, т. к. эта сумма может быть то большей, то меньшей. Н. Е. Введенский объяснил это явление с точки зрения состояния возбудимости мышцы, введя понятие об оптимуме и пессимуме частоты раздражения.
Оптимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу повышенной возбудимости. Тетанус при этом будет максимальным по амплитуде - оптимальным.
Пессимальной называется такая частота раздражения, при которой каждое последующее раздражение осуществляется в фазу пониженной возбудимости. Тетанус при этом будет минимальным по амплитуде - пессимальным.
Режимы мышечных сокращений. Различают изотонический, изометрический и смешанный режимы сокращения мышц.
При изотоническом сокращении мышцы происходит изменение ее длины, а напряжение остается постоянным. Такое сокращение происходит в том случае, когда мышца не перемещает груз. В естественных условиях близкими к изотоническому типу сокращений являются сокращения мышц языка.
При изометрическом сокращении длина мышечных волокон остается постоянной, меняется напряжение мышцы. Такое сокращение мышцы можно получить при попытке поднять непосильный груз.
В целом организме сокращения мышц никогда не бывают чисто изотоническим или изометрическим, они всегда имеют смешанный характер, т. е. происходит изменение и длины, и напряжения мышцы. Такой режим сокращения называется ауксотоническим если преобладает напряжение мышцы, или ауксометрическим если преобладает укорочение.
Механизм мышечного сокращения. Мышцы состоят из мышечных волокон, которые состоят из множества тонких нитей - миофибрилл, расположенных продольно. Каждая миофибрилла состоит из протофибрилл - нитей сократительных белков актина и миозина. Перегородки, называемые 2-пластинами, разделяют миофибриллы и, следовательно, мышечное волокно на участки - саркомеры. В саркомере наблюдают правильно чередующиеся поперечные светлые и темные полосы. Эта поперечная исчерченность миофибрилл обусловлена определенным расположением нитей актина и миозина. В центральной части каждого саркомера свободно расположены толстые нити миозина. На обоих концах саркомера находятся тонкие нити актина, прикрепленные к Z-пластинам. Нити миозина выглядят в световом микроскопе как светлая полоска (Н-зона) в темном диске, который дает двойное лучепреломление, т. к. содержит нити миозина и актина и называется анизотропным или А-диском. По обестороны от А-диска находятся участки, которые содержат только тонкие нити актина и кажутся светлыми, т. к. они обладают одним лучепреломлением и называются изотропными или j -дисками. По их середине проходит темная линия - Z-мембрана. Именно благодаря такому периодическому чередованию светлых и темных дисков сердечная и скелетная мышцы выглядят исчерченными (поперечно-полосатыми) (рис. 6).
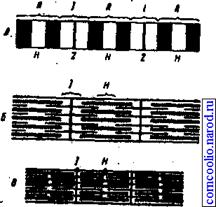
Рис. 6. Электронномикроскопическая картина миофибриллы (схематизировано)(А). Взаимное расположение толстых (миозиновых) и тонких (актиновых) нитей в расслабленной (Б) и сокращенной (В) миофибрилле.
В состоянии покоя концы толстых и тонких: нитей лишь незначительно перекрываются на уровне А-диска. В соответствии с теорией скользящих нитей при сокращении тонкие актиновые нити скользят вдоль толстых миозиновых нитей, двигаясь между ними к середине саркомера. Сами актиновые и миозиновые нити своей длины не изменяют.
Механизм скольжения нитей. Миозиновые нити имеют поперечные мостики (выступы) с головками, которые отходят от нити биполярно. Актиновая нить состоит из двух закрученных одна вокруг другой цепочек (подобно скрученным ниткам бус) молекул актина. На нитях актина расположены молекулы тропонина, а в желобках между двумя нитями актина лежат нити тропомиозина. Молекулы тропомиозина в покое располагаются так, что предотвращают прикрепление поперечных мостиков миозина к актиновым нитям.
Во многих местах участки поверхностной мембраны мышечной клетки углубляются в виде трубочек внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-систему). Параллельно миофибриллам и перпендикулярно поперечным трубочкам расположена система продольных трубочек (альфа-система). Пузырьки на концах этих трубочек - терминальные цистерны - подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В этих пузырьках сосредоточено основное количество внутриклеточного кальция.
В состоянии покоя миозиновый мостик заряжен энергией (миозин фосфорилирован), но он не может соединиться с нитью актина, так как между ними находится система из нитей тропомиозина и глобул тропонина. При возбуждении ПД быстро распространяется по мембранам поперечной системы внутрь клетки и вызывает высвобождение ионов кальция из альфа-системы. С появлением ионов кальция в присутствии АТФ происходит изменение пространственного положения тропонина, в результате чего отодвигается нить тропомиозина и открываются участки актина, присоединяющие ми-озиновые головки. Соединение головки фосфорилированного миозина с актином приводит к изменению положения мостика (его "сгибанию"), в результате конформации этой части миозиновой молекулы, и перемещению нити актина на один шаг (на один "гребок") к середине саркомера. Затем происходит отсоединение мостика от актина. Ритмические прикрепления и отсоединения головок миозина позволяют "грести" или тянуть актиновую нить к середине саркомера.
При отсутствии повторного возбуждения ионы кальция закачиваются кальциевым насосом из протофибриллярного пространства в систему саркоплазматического ретикулума. Это приводит к снижению концентрации ионов кальция и отсоединению его от тропонина. Вследствие чего тропомиозин возвращается на прежнее место и снова блокирует активные центры актина. Вместе с тем, происходит фосфорилирование миозина за счет АТФ, который не только заряжает системы для дальнейшей работы, но и способствует временному разобщению нитей. Удлинение (расслабление) мышцы после ее сокращения является процессом пассивным, поскольку актиновые и миозиновые нити легко скользят в обратном направлении под влиянием сил упругости мышечных волокон и мышцы, а также силы растяжения мышц антагонистов.
Гладкие мышцы. Гладкие мышцы, формирующие мышечные слои стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других полых внутренних органов, построены из веретенообразных одноядерных мышечных клеток. Отдельные клетки в гладких мышцах функционально связаны между собой низкоомны-ми электрическими контактами - нексусами. За счет этих контактов потенциалы действия и медленные волны деполяризации беспрепятственно распространяются с одного мышечного волокна на другое. Поэтому несмотря на то, что двигательные нервные окончания расположены на небольшом числе мышечных волокон, вследствие беспрепятственного распространения возбуждения с одного волокна на другое в реакцию вовлекается вся мышца. Следовательно, гладкие мышцы представляют собой не морфологический, а функциональный синцитий.
Особенностью гладких мышц является их способность осуществлять относительно медленные движения и длительные тонические сокращения. Медленные, имеющие ритмический характер, сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц особенно хорошо выражены в сфинктерах полых органов, которые препятствуют выходу содержимого этих органов.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов.
Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. Эти различия хорошо наблюдать при медленном растяжении гладкой и скелетной мышцы. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеет большое значение для нормального функционирования полых органов. Благодаря высокой пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии. Так, например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления внутри его.
Сильное и резкое растяжение гладких мышц вызывает их сокращение. Последнее обусловлено нарастающей при растяжении деполяризацией клеток, обусловливающих автоматию гладкой мышцы. Сокращение, индуцируемое растяжением, играет важную роль в авторегуляции тонуса кровеносных сосудов, а также обеспечивает непроизвольное (автоматическое) опорожнение переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
В гладких мышцах одиночное сокращение продолжается несколько секунд. Тетаническое сокращение возникает при низкой частоте слияния одиночных сокращений и низкой частоте сопровождающих его ПД.
В отличие от скелетной мышцы гладкая мышца кишки, мочеточника, желудка и матки развивает спонтанные тетанообразные сокращения в условиях ее изоляции и денервации, и даже после блокады нейронов интрамуральных ганглиев. В этом случае они возникают не в результате передачи нервных импульсов с нерва, а вследствие активности клеток, обладающих автоматией, т. е. пейсмекерных клеток. Последние идентичны по структуре другим мышечным клеткам, но отличаются по электрофизиологическим свойствам. В этих клетках возникают препотенциалы или пейсмекерные потенциалы, депо-ляризующие мембрану до критического уровня. Вследствие входа, главным образом, ионов кальция мембрана деполяризуется до изо-электрического уровня, а затем поляризуется с обратным знаком (до + 20 мВ). Реверсия потенциала длится в течении нескольких секунд. За реполяризацией следует новый препотенциал, который вызывает еще один потенциал действия.
Вегетативная нервная система и ее медиаторы оказывают на спонтанную активность пейсмекеров модулирующие влияния. При нанесении ацетил холина на препарат мышцы толстой кишки пейсмекерные клетки деполяризуются до околопорогового уровня и ча-стота ПД возрастает. Инициируемые ими сокращения сливаются, образуется почти гладкий тетанус. Чем выше частота ПД, тем сильнее суммированное сокращение. Нанесение на этот препарат норадрена-лина гиперполяризует мембрану и таким образом снижает частоту ПД и величину тонуса.
Возбуждение гладкомышечных клеток вызывает либо увеличение входа ионов кальция через мембрану клетки, либо высвобождение ионов кальция из внутриклеточных хранилищ. В результате повышения концентрации ионов кальция в саркоплазме активируются сократительные структуры. Так же как сердечная и скелетная мышца, гладкая мышца всегда пассивно расслабляется, если концентрация ионов кальция очень мала. Однако расслабление гладких мышц происходит более медленно, т. к. замедлено удаление ионов кальция.








