 2015-04-08
2015-04-08 2016
2016Несмотря на то, что некоторое количество нуклеозидов и свободных азотистых оснований образуется при гидролизе полинуклеотидов пищи, большая часть включающихся в нуклеиновые кислоты нуклеотидов синтезируется заново.
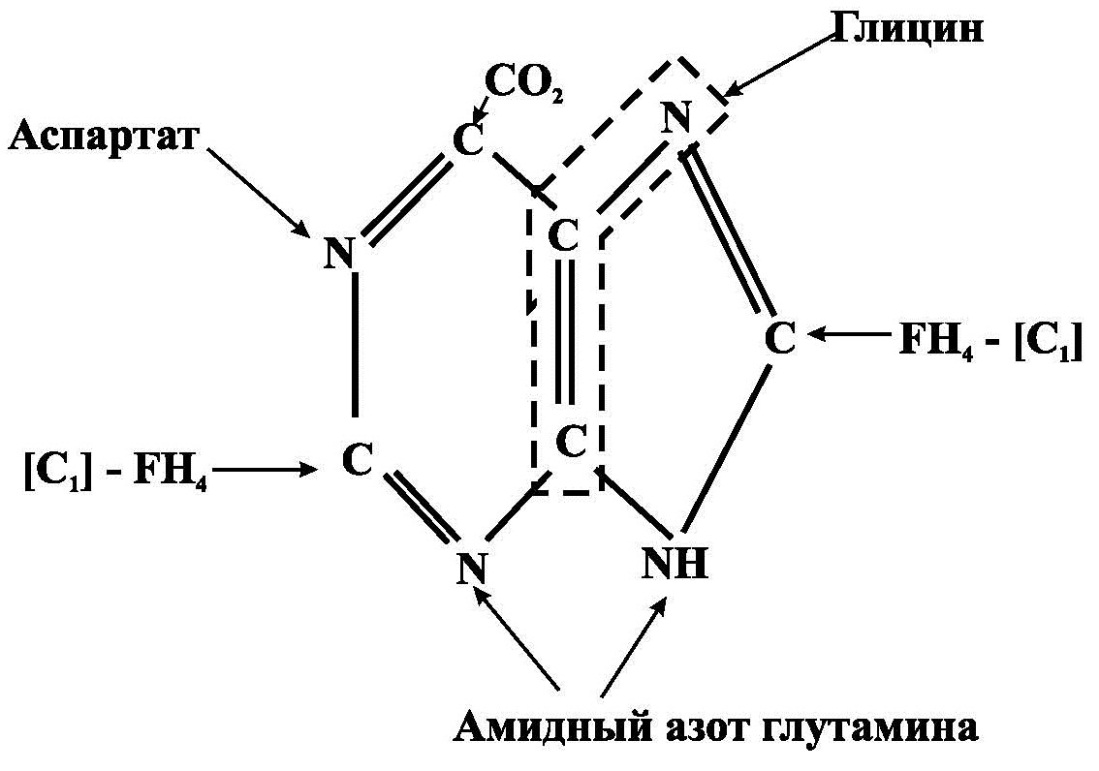
Рис.1. Предшественники пуринового кольца
Синтез пуриновых нуклеотидов начинается с того, что к рибозо-5- фосфату поэтапно присоединяются различные соединения, из которых формируется пуриновое кольцо. Предшественниками пуринового кольца являются: аспартат, глутамин, СО2, глицин, кроме того два атома углерода поставляет тетрагидрофолат (рис.1). Первым синтезированным пуриновым нуклеотидом является инозинмонофосфат (ИМФ), из него затем образуются АМФ и ГМФ. Печень является основным местом образования пуриновых нуклеотидов. Она снабжает пуринами ткани, не способные к их синтезу: клетки крови и частично мозг.
Упиримидиновых нуклеотидов (в отличиеот пуриновых) сначала из предшественников собирается пиримидиновое кольцо, а затем к готовому кольцу присоединяется рибозо-5-фосфат. Источниками пиримидинового кольца являются аспартат и карбамоилфосфат. Первым синтезированным пиримидиновым нуклеотидом является оротидинмонофосфат (ОМФ). Из него при декарбоксилировании образуется УМФ; из УТФ при воздействии глутамина – ЦТФ. Для синтеза ТМФ из дУМФ необходим тетрагидрофолат.
Для синтеза нуклеиновых кислот необходимы нуклеозидтрифосфаты (НТФ), которые образуются из нуклеозидмонофосфатов (НМФ) при участии АТФ: НМФ"НТФ. Для синтеза ДНК необходимы дезоксиНТФ, которые образуются путем восстановления нуклеозиддифосфатов (НДФ) в дезоксиНДФ при участии фермента рибонуклеотидредуктазы(рис.2). Непосредственным восстановителем НДФ является глутатион или белок тиоредоксин, сульфгидрильные группы которых в ходе этой реакции окисляются.
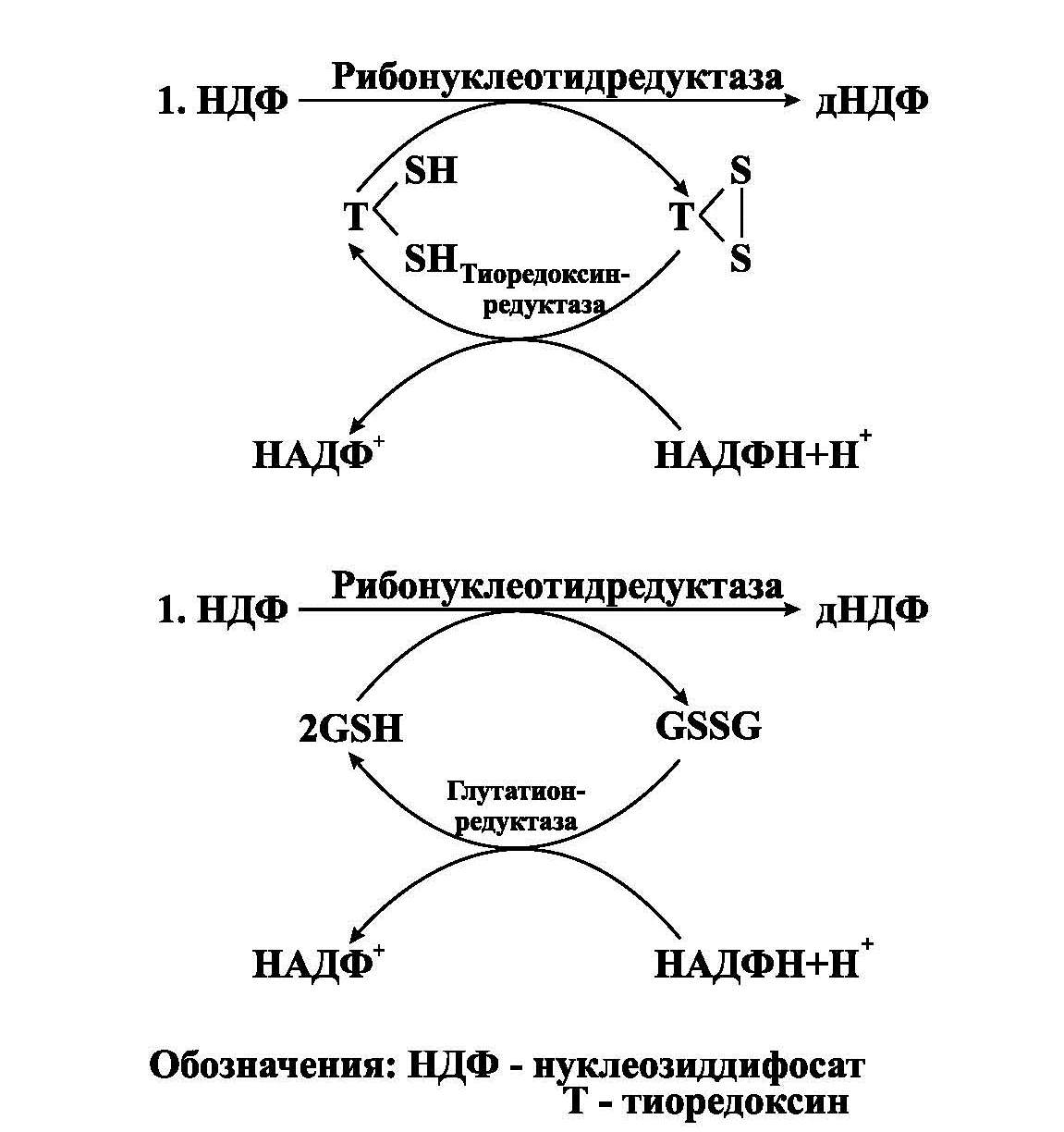
Рис.2. Синтез дезоксирибонуклеотидов
Катаболизм пуриновых нуклеотидов происходит без разрыва пуринового кольца и приводит к образованию мочевой кислоты, которая плохо растворима (рис.3).При повышении концентрации мочевой кислоты в плазме крови возникает гиперурикемия, которая может явиться причиной развития подагры.
При подагре кристаллы мочевой кислоты и уратов откладываются в суставных хрящах, являясь причиной острого воспаления суставов (чаще всего мелких). Кроме того, ураты могут образовывать уратные камни, что может статьпричиной почечной недостаточности. Характерным для подагры симптомом является и отложение уратов в подкожной клетчатке с образованием тофусов.
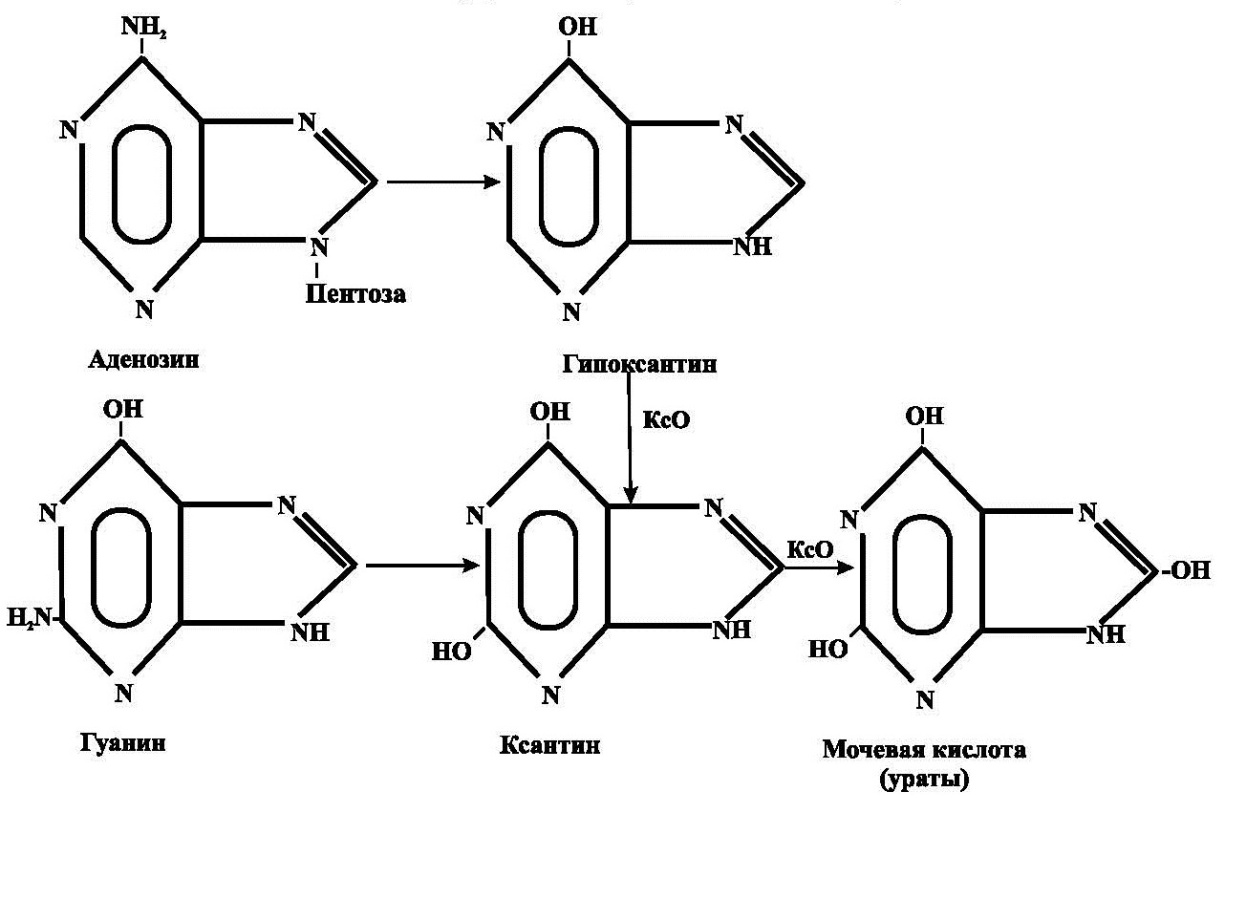
Рис.3. Катаболизм пуринов
Катаболизм пиримидинов (рис.4.) происходит с разрывом пиримидинового кольца, конечными продуктами его являются аммиак, СО2, β-аланин (образуется при катаболизме урацила и цитозина) и β-аминоизомасляная кислота (образуется при катаболизме тимина).
Нуклеиновые кислоты представляют собой полинуклеотиды. ДНК и РНК содержат разный набор нуклеотидов. ДНК содержит тимин, цитозин, гуанин, аденин, РНК вместо тимина содержит урацил. В образовании РНК принимают участие рибонуклеотиды, а ДНК -дезоксирибонуклеотиды. Функции, выполняемые разными видами нуклеиновых кислот, также различны: все виды РНК участвуют в биосинтезе белка, а ДНК участвует в сохранении, передаче наследственной информации и делении клетки. Первичная структура нуклеиновых кислот представляет собой определенную последовательность НМФ, соединенных фосфодиэфирными связями. Вторичная структура РНК представляет одиночную спираль с участками удвоения, а вторичная структура ДНК представляет двойную спираль (дуплекс), у которой цепи антипараллельны и комплементарны на всемпротяжении, т.е. напротив аденина (А) одной цепи стоит тимин (Т) в другой цепи,напротив гуанина (Г) стоит цитозин (Ц).
Поэтому, согласно правилу Чаргаффа, число пуринов (А+Г) равно числу пиримидинов (Т+Ц). Между комплементарными азотистыми основаниями возникают водородные связи, которые удерживают обе цепи дуплекса (между А и Т две связи, между Г и Ц– три). Комплементарные основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль.
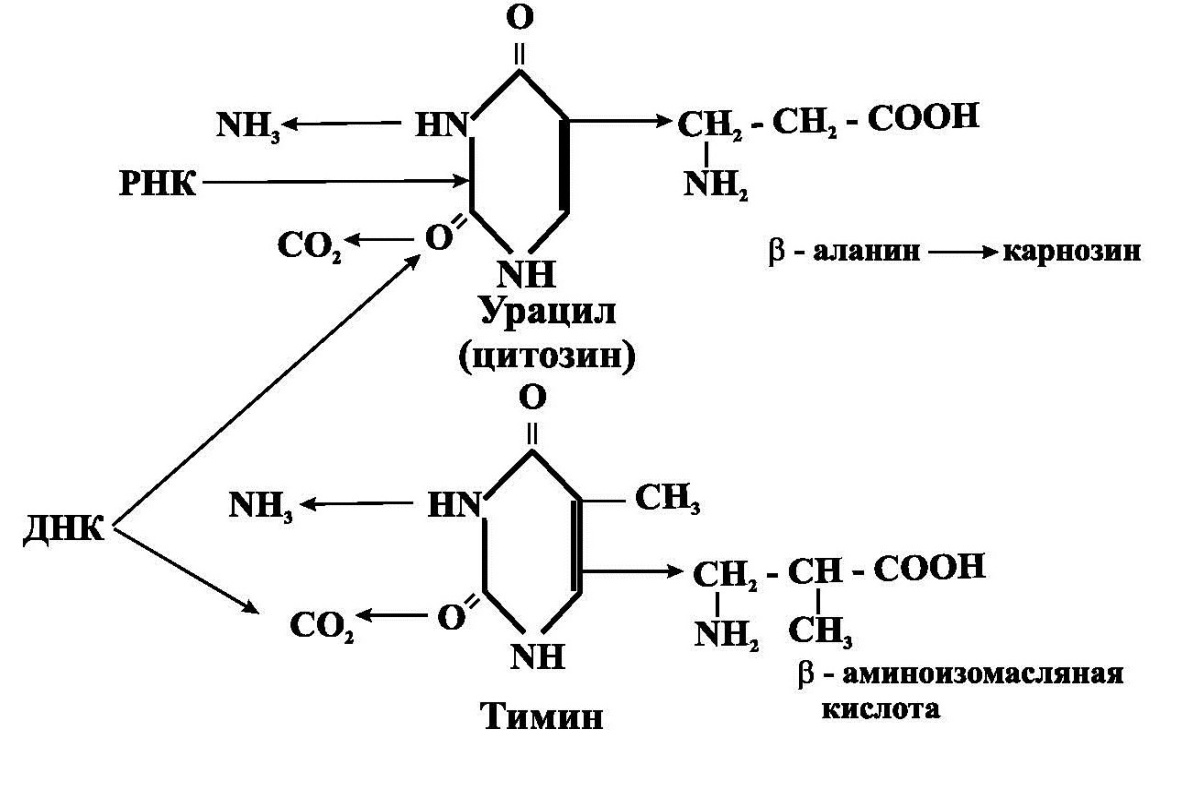
Рис.4. Катаболизм пиримидинов
Третичная структура ДНК представляет собой суперспираль, благодаря которой длина ДНК укорачивается в 10000 раз. Суперспирализация осуществляется с помощью разнообразных белков, т.е. с химической точки зрения это нуклеопротеин (комплекс белков и ядерной ДНК называют хроматином). Структурной единицей хроматина являются нуклеосомы, которые образуются путем «накручивания» молекулы ДНК вокруг «нуклеосомного кора», состоящего из 8 молекул гистоновых белков типа Н2- Н4.
В клетке существуют разные виды РНК, которые выполняют разные функции. Рибосомальная РНК (рРНК) участвует в образовании рибосом, на которых происходит биосинтез белка. Матричная РНК (мРНК) передает информацию с ДНК на синтезированный белок, транспортная РНК (тРНК) участвует в активации аминокислот и передает их на рибосомы. Малые некодирующие РНК (мнкРНК) выполняют каталитическую и регуляторную функции.
В основе всех матричных биосинтезов лежит принцип комплементарности. Для матричных биосинтезов необходима матрица (нуклеиновая кислота), с которой будет считываться информация; активированные мономеры или субстраты для синтезов (НТФ или аминоацил-тРНК), катализаторы (в их роли выступают ферменты и рибозимы), а также различные белковые факторы матричных биосинтезов.

