 2015-04-12
2015-04-12 4716
4716Процесс раскручивания двойной спирали в репликативной вилке порождает механические и топологические проблемы. В принципе раскручивание линейной двунитевой ДНК может происходить благодаря вращению родительской спирали вокруг собственной оси Но такое вращение очень длинных цепей ДНК вокруг длинных же осей во внутриклеточном пространстве механически затруднено. При репликации замкнутых кольцевых ДНК раскручивание цепей в вилке создает дополнительные проблемы, По мере раскручивания цепей степень отрицательной сиерхспиральности сегментов, находящихся перед вилкой репликации, постепенно уменьшается и в них возникает положительная сверхспирализация. Дальнейшее перемещение вилки вдоль кольца затрудняется и в конце концов блокируется. Это блокирование снимается путем внесения однонитевого или двунитевого разрыва. Тем самым образуется «шарнир», который дает возможность нереплицированному дуплексу, находящемуся перед вилкой, вращаться вместе с ней. Такие разрывы вносятся в ДНК с помощью ферментов, имеющих общее название ДНК-топоизомеразы. Эти ферменты изменяют степень сиерхспиральности и тип сверхспирали. В различных организмах идентифицированы топоизомеразы двух основных типов. Одни ферменты, называемые топоизомеразами типа I надрезают одну из двух нитей, в результате чего фланкирующие дуплексные области могут повернуться вокруг интактной цепи, и затем воссоединяют концы разрезанной цепи, как показано на рис…. Эта реакция не требует энергии АТР,поскольку энергия фосфодиэфирной связи сохраняется благодаря тому, что тирозиновый остаток в молекуле фермента выступает то в роли акцептора, то в роли донора фосфорильного конца разрезанной нити.
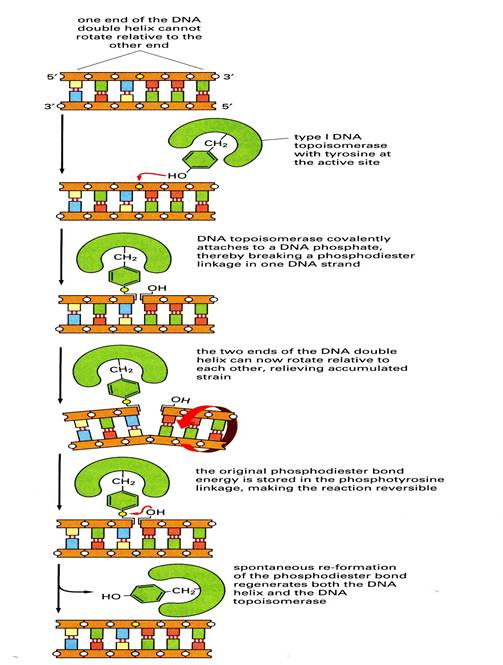
Рис… Принцип действия топоизомеразы I
Отмечены два интересных, но, возможно, не связанных друг с другом различия между топоизомеразами I типа у про- и эукарисп: во-первых, топоизомеразы I прокариот взаимодействуют с 5'-фрсфорильным концом разорванной цепи, а эукариот- с 3'-фосфорилыным концом, а во-вторых топоизомеразы I прокариот устраняют только отрицательные сверхвитки, а ‘укариотические - как отрицательные, так и положительные.
Топоизомеразы II типа устраняют как отрицательные, так и положительные сверхвитки. В отличие от топоизомераз I типа топоизомеразы II вносят временные разрывы в обе комплементарные нити, пропускают двунитевой сегмент той же самой или другой молекулы ДНК через разрыв, а затем соединяют разорванные концы, как это показано на рис….. Топоизомеразы II типа тоже используют гирозиновые остатки (присутствующие по одному в каждой из субъединиц ферментов) для связывания 5'-конца каждой разорванной нити в то время, когда другой дуплекс проходит через место разрыва. В результате внесения двухцепочечного разрыва и прохождения через него другого дуплекса за одну реакцию снимаются два отрицательных или положительных сверхвитка. В некоторых случаях дуплексом, проходящим через место разрыва, оказывается другая замкнутая молекула ДНК; это приводит к разделению сцепленных кольцевых ДНК или, напротив, к образованию таких сцепленных комплексов (катенанов). Этот механизм может использоваться и для распутывания или запутывания клубков, а также для раскручивания или конденсации крупных дуплексных ДНК.
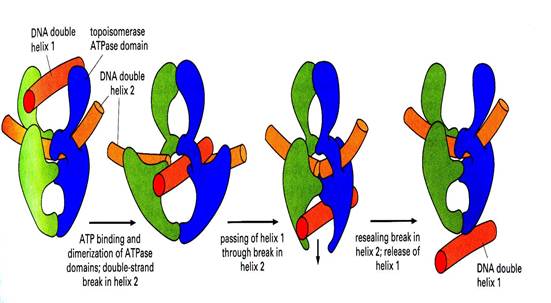
Рис… Принцип работы топоизомеразы II
Топоизомеразы типов I и II снимают сверхвитки, образующиеся при репликации кольцевой ДНК. Однако существует особая топоизомераза II, называемая гиразой и обнаруженная пока только у бактерий, которая индуцирует образование отрицательных свсрхвитков в релансированных кольцевых ДНК. Для этого гираза делает двунитевые надрезы и затем особым способом воссоединяет концы, таким образом снимая положительные сверхвитки и внося отрицательные в релаксировашгую ДНК.
У бактерий топоизомераза I и гираза являются ключевыми ферментами, определяющими степень суперскрученности ДНК при ее ответе на стрессовые внешние воздействия – такие как повышение температуры, изменении рН и оксидативный стресс. Сбалансированное действие топоизомеразы I и гиразы у бактерий регулирует степень сверхспиральности ДНК и влияет на скорость движения репликативной вилки.
У эукариот ТорII является одним из компонентов клеточного ответа на различные типы повреждений ДНК. Она образует комплекс с важнейшими регуляторами клеточного цикла, включая Р53, BRCT-содержащий белок TopBP1 и Cdc2-киназу. Ассоциация топоизомеразы II с Cdc2-киназой указывает на начальную раннюю стадию конденсации хроматина в митозе. Регуляция этого взаимодействия может являться новым механизмом, влияющим на вступление клеток в митоз. Конденсация хромосом и их сегрегация при переходе в анафазу находятся под контролем белкового комплекса, состоящего из топоизомеразы II и конденсина.
В кроссоверном разрешении холидеевских структур могут принимать участие различные эндонуклеазы, в том числе XPF. При некроссоверном разрешении, при котором также нужны топологические изменения ДНК, их могут проводить RecQ-подобные геликазы вместе с топоизомеразой III. Она также, как и топоизомераза II способна генерировать и воссоединять двунитевые разрывы ДНК.
RecQ-подобные геликазы вместе с топоизомеразой III играют важную роль не только в гомологической рекомбинации, но и в других клетчных процессах, таких как репликация и контроль клеточного цикла.
Терминация репликации.








