 2015-04-06
2015-04-06 3450
3450Ретинолдың өнеркәсіптік өндірісі микробиологиялық және химиялық синтезбен өтеді.
Микробиологиялық синтезі B. trispora қолдануына негізделген, субстрат ретінде бидай немесе күріш ұны, өсімдік майы. Бета-каротин синтезі стимуляторлары – В-ионон немесе цитрустық меласса және тиамин. Күн сәулесі осы пигменттің шығуын күшейтеді.
B. trispora (+) және (-) штамдары бөлек қоректік орталарға егіледі, ферментация алдында оларды биореакторда араластырады. (+) және (-) штамдардың қатынасы 1/15 тен, яғни еркек формасы (-) 15-еселік шектен тыс көп болуы. Әр түрлі жынысты аналық (+) және аталық (-) мицелийлері біріккенде зигота пайда болады, ол жеке штамдарға қарағанда 5-17 есе жоғары бета-каротинді түзеді. Ферментация аэрация мен тұрақты араластыруымен жүреді. Бета-каротинге активті ауысуынан майлардың концентрациясы төмендейді. Бета-каротиннің деңгейі – 2000 мг/л жетеді.
Ферментация аяғанда биомассаны сепаратордан өткізеді, аэрозольді кептіру жүргізеді, қалдық ылғалы 7% дейін болады. Қолдану мақсатына байланысты келесі технологияларды ажыратады:
- кептіргеннен кейін ұнтақты толтырғышпен араластырады және грануллайды, жем концентраты ретінде қолданады;
- маймен экстракциялайды, концентрациялайды, этанолмен шаяды, каротинді майдан алады (күнбағыс немесе басқасы), онда провитаминнің деңгейі 2,0-2,5 г/кг, нанға тағам қоспасы ретінде пайдаланады;
- Кристалды бета-каротин-фреонмен экстракциялайды, органикалық қоспалардан тазартады, қызғылтсары түсті кристалдар алады, медициналық мақсатта қолданады.
Эргостериннің алу көзі (эргоста-5,7,22-триен-3β-ол) микробалдырлар, зең саңырауқұлақтары әне оларға аса бай ашытқылар. Сондықтан витамин D алу үшін алғашында микробиологиялық синтезімен ашытқылардың биомассасын жинайды. Кейін ашытқы суспензиясын немесе кептірілген ашытқы клеткаларын ультракүлгін сәулесімен өндейді, нәтижесінде эргостериннің фотохимиялық алмасуында эргокальциферол пайда болады (57-сурет).
Эргостеринді өндірушілер қатарында Saccharamyces carisbergensis, S.cerevisiae, S.ellipsides және басқа микроорганизмдер қолданылады
(10-кесте). Эрго-стерин бактерия клеткаларында өте аз мөлшерде түзіледі.
Эргостериннің бастауышы-сквален. Ашытқы клеткаларында сквален синтезі дақыл қартая бастағанда күшейеді және өсудің стационарлық фазасында жалғасады.
Ашытқылардың ферментациясы аэрация жағдайында және азот көзімен салыстырғанда көмірсулардың көздерінің шектен тыс асқанда іске асады. Алынған биомассаны тұз қышқылы ерітіндісімен гидролиздейді, кейін спиртпен тазалайды, концентрациялайды, 280-300 нм толқындар ұзындығымен ультракүлгін сәулесімен өңдейді. Осындай сәулелер А және В көміртектік циклдарда химиялық байланыстарды қоздырады, нәтижесінде эргостерин D витаминге айналады.
Витаминнің майлы формасын алу өнімді фильтрациядан өткізгеннен кейін өсімдік маймен ерітеді және тағамдық қоспа ретінде пайдаланылады.
Кристаллды витамин дайындау үшін қосымша тазарту жүргізіледі.
Микроорганизмдерде эргостерин мөлшері
| Микроорганиздер | Эргостерин мөлшері,% (құрғақ затқа) |
| Saccharomyces carlbergensis | 0.49-4.3 |
| Saccharomyces ellipsodes | 1.2-1.5 |
| Rhodotorula glutinis | 0.7-0.9 |
| Candida utilis | 0.4-0.6 |
| Candida tropicalis | 0.2-0.3 |
| Aspergilus niger | 1.2-1.4 |
| Penicillium westlingii | 2.2 |
5.Гаплоидтерді алу тәcілдері. Табиатта гаплоидтер апогамия, гиногенез немесе андрогенез нтижесінде мейоздың редукциялыќ бөліну барысында аталыќ немесе аналыќ жыныс жасушаларынан пайда болады. Редукцияланған апогамия - ўрыќтың синергидтер немесе антиподтардан пайда болуы
Гиногенез /аналыќ партеногенез/ - спермий тозаңдану кезінде жўмыртќа жасушасына енген соң јрі ќарай дамымайды, нәтижесінде ўрыќтануға дайын, біраќ ўрыќтанбаған жўмыртќа жасушасы гаплоидтік ўрыќты тїзеді.
Андрогенез /аталыќ партеногенез/ - спермий жўмыртќа жасушасына еніп, оныѕ ядросын жарамсыз етеді. Спермий ядросы жўмыртќа цитоплазмасымен бірге бөліне бастайды, оныѕ нәтижесінде аталыќ хромосомалар жиынтыєы бар гаплоидтік ўрыќ пайда болады.
Өсімдіктерді химиялыќ заттармен, иондаушы сәулемен және температураны күрт өзгертіп өңдеу, сәулелендірілген тозаңмен тозаңдандыру, алшаќ будандастыруды ќолдану – осының бірі гаплоид алу үшін жасалғанымен, оның пайда болу дәрежесі өте төмен. Сондыќтан бўл тәсілдер гаплоидті тўраќты түрде алуга мүмкіндік бермейді.
Ќазіргі кезде гаплоидтерді алу үшін мынандай тәсілдерді ќолданады:
1) Псевдогамия - жўмыртќа жасушасын бөтен тозаңмен тозаңдандырған соң ол ўрыќтанбайды, оның нәтижесінде гаплоидтік ўрыќ пайда болады.
2) Гаплопродюсер тәсілі - будан ўрығында атаґаналар хромосомаларының бір жиынтығы іріктеліп жойылады.
3) Андрогенез және гиногенез in vitro тјсілдері - микроспоралар мен тозаңќапты (андрогенез), тўќымбүршік пен ўрыќ ќалтасын (гиногенез) ќоректік ортада өсіру арќылы гаплоидті өсімдігін алу.
Псевдогамия барысында гаплоидтердің пайда болуы
Түраралыќ будандастырудың кейбір тіркесімдері ќўйылысу барысында ўрыќ пен тўќым ўрыќтанбаған аналыќ гаметадан түзілуі мїмкін.
Сонымен ќатар псевдогамия сәулемен өңделген тозаңды ќолданғанда немесе әр түрлі плоидтыќ деңгейдегі түрлерді будандастырғанда жүзеге асады.
Гиногенезге ұшырататын будандастырудың нәтижесінде түзілетін ўрыќтанбаған ќызылша тўќымбүршіктерін зерттеген кезде, гиногенетикалыќ ўрыќтардың жўмыртќа жасушасынан немесе антиподтан дамитындығы аныќталды. Синергидтер көбінесе жойылады. Өсірілген 1000 тўќымбүршіктен 2 гаплоидті өсімдік алуға болады. Колхицинмен өңдеу арќылы гомозиготалыќ дигаплоидті өсімдік алынып, танаптыќ жағдайында зерттелді.
Гаплопродюсер тјсілі (Бульбозум тјсілі). Кейбір алшаќ түрлерді будандастырған кезде ўрыќтың алғашќы даму кезінде ата-анасыныѕ біреуінің хромосомалар жиынтығы жойылатындығы аныќталды. Бўл тәсіл селекция жўмысында арпаның жабайы Hordeum bulbosum түрімен будандастыруда кеңінен пайдаланылады. Hordeum vulgare жўмыртќа жасушасын Hordeum bulbosum тозаѕымен ўрыќтандырғанда генетикалыќ сәйкессіздікке байланысты аталыќ хромосомасы жоғалады да, гаплоидті ўрыќ дами бастайды. Оны зиготаныѕ алєашќы бөліну кезінде байќауға болады. Бўл Hordeum bulbosum геномында хромосоманың митотикалыќ ўршыќ жіпшелеріне жалғасу механизмнің болмауы деген болжау бар. Аналыќ өсімдікте ўрыќтың ќалыпты өсуі тежеледі, сондыќтан оны оќшаулап, ќоректік ортада өсіріп жетілдіреді. Одан кейін гаплоидті өскіндерді хромосома санын екі еселеу және гомозиготалыќ изогендік линиялар алу їшін колхицинмен өңдейді.
Осы тәсілмен Канадада арпаның Минго және Родео сорттары шығарылды.
Бүкілодаќтыќ селекция, генетика институтында гаплоидтік селекция негізінде, ќысќа мерзім ішінде (8-10 жыл орнына 4-5 жыл ішінде) арпаның Исток, Одесский-15 сорттары шығарылды. Олар жоғары өнімді, жатып ќалуєа және ќаракүйеге төзімді. Элита Поволжья деп аталатын ғылыми кәсіподаќ осы тјсілдерді ќолдана отырып, ќуаңшылыќќа төзімді 700-ден астам арпа линияларын алды. Оңтүстік-Шығыс ауылшаруашылыќ ғылыми–зерттеу институтында көптеген жаздыќ бидай мен арпа линиялары алынды. Бүкілодаќтыќ ауылшаруашылыќ биотехнологиясының ғылыми-зерттеу институтында (НИИСХБ) болашағы бар бидайдың андрогендік линиялары алынды.
In vitro жағдайында ќоздырылған андрогенез
Жекелеген тозаңќаптарды өсірген кезде олардың тозаңдарының ќалыпты гаметофит арќылы даму жолы тежеліп, спорофиттік жолға көшкен кезде гаплоидтік өсімдік пайда болады. Ќазіргі кезде осы тәсіл бойынша 200-ден астам өсімдік түрі алынды. Әсіресе, бўл тәсілмен Ќытай селекционерлері нәтижелі жўмыс істеді. Олар жаңа жоғары өнімді ірі төзімді күріш пен бидай сорттарын шығарды. Бүгінгі таңда сол күріш 170000 га, бидай 70000 га алќапта өсіріледі. 13-суретте тозаңќап өсіндісі көрсетілген
Тозаѕќапты өсіру. Ќазіргі кезде тозаңќаптан гаплоидті өсімдікті алу әдісі кеңінен ќолданылып келеді. Тозаңќап өсіндісінде микроспорадан эмбриоид ќўрылымы немесе каллус ўлпасы тїзіледі. Эмбриоид ќўрылымын арнайы жағдайда өсірген кезде хромосомаларының жиынтығы гаплоидты өскін алуға болады. Мўны тікелей андрогенез дейді. эмбриоидтер тозаңќапты жарып, микроспорадан тікелей дамиды.
Көп жағдайда регенеранттар каллусогенез арќылы пайда болады /мысалы, күріште/, мўны жанама андрогенез дейді. Егер микроспорадан каллус ўлпасы ќалыптасса (15-сурет), гаплоидті өсімдік алу күрделене түседі, себебі алдымен сабаќ органогенезін ќоздыру керек, кейін одан өркен түзілуі жылдамдатылады, бўл әрќашан бола бермейтін жағдай.
Сонымен ќатар, каллус ўлпасы генетикалыќ тўраќтылыќќа ие болмайды, одан алынған регенеранттар алғашќы өсімдіктегі бағалы ќасиетті саќтай алмайды, кейде одан пайда болған өсімдіктер гаплоидті болмауы мүмкін. Сондыќтан тіршілікке икемді эмбриоидтерді микроспорадан алу өте сирек болғанымен, генетикалыќ және селекциялыќ жўмыстар үшін гаплоидтерді алу їшін бірінші тәсіл-тікелей андрогенез оңтайлы болып отыр.
Тозаңќапты өсіру арќылы гаплоидтерді алу тәсілінің мәні мынада. Алдын ала залалсыздандырылған бітеугүлден in vitro жағдайда тозаңќаптарды айырады. Оларды ќолма-ќол ќоректік ортаға орналастырады, орта ќўрамын әрбір өсімдікке арнайы таңдап дайындайды. Тозаңќапты алу сәтінде олардың микроспоралары эмбриогенезге көшу үшін ќолайлы даму кезеңінде болғаны өте маңызды.
Көптеген жағдайда тозаңќапты айыру үшін тиімді кезеңі деп микроспораның ортаңғы немесе соңғы даму кезеңін айтады. Бўл кезде микроспоралар тетрададан бөлініп, бірінші митозға дайындалады. Біраќ көптеген өсімдік түрлері үшін тетрада кезеңі мен екі ядролыќ тозаң түйіршігі кезеңі аралығында тиімді болу мүмкін. Мысалы, ќырыќќабаттар үшін тиімді кезең микроспораның бір ядролыќ алғашќы даму кезңі болса, бидай микроспоралары үшін бір ядролыќ ортаңғы даму кезеңі болып саналады.
Микроспоралар ќалыпты дамуынан ауытќу нәтижесінде тікелейандрогенезге өтуі мүмкін, бўл жағдайда гаплоидті ўрыќ, каллус түзбей аќ, тікелей микроспорадан /вегетативтік немесе генеративтік жасушадан/ пайда болады немесе ол каллус түзе бастайды. Бірінші тікелей андрогенез жағдайында микроспора бөлінеді де, діңґєќ жасушалыќ проэмбрио түзеді, ўрыќ глобула тјріздес кезеңінде экзинаны жарып, әрі ќарай дамиды.
Каллус ўлпасы түзілген /тікелей жол емес/ жағдайда тозаң бөлінеді, біраќ бөліну кезінде пайда болған жасушалар тез ўлғаяды жјне тозаң түйіршігінің ќабығын жарып, каллус массасын ќўрайды. Микроспоралардан гаплоид регенераттары түрлі жолдармен пайда болады (17- сурет, ќосымша А).
Тозаңќаптан алынған өсімдіктің ерекшелігі негізінен оның тікелей андрогенез немесе каллус ўлпасы арќылы пайда болғандығына байланысты. Тікелей андрогенез жолымен алынса, нағыз гаплоидті өсімдік шығады -1n, ал каллустан өсірілгенде, ди, три, тетраплоидтер, яғни 2n,3n,4n, жјне т.б. эуплоидтер, анеуплоидтер, сонымен ќатар, басќа генетикалыќ өзгерістері бар өскіндер алынады (19-сурет).
Жасушалардың полиплоидтылыєы ядролардың ќосылуы, хромосомалардың эндоредупликациясы, сандарының өзгеруі және микроспораныѕ түзілуіне байланысты болуы мүмкін. Бўл жағдайда гаметоклоналдыќ варианттар деп аталатын өсімдіктер ќалыптасады.
I-эмбриоидоген
Гаметоклоналдыќ өзгергіштік-микроспора, тозаңќап, ўрыќ ќалтасын, яғни гаметаларды ќоректік ортада өсіргенде жүзеге асатын өзгергіштік.
Жасуша селекциясындаєы сияќты, микроспора өсіндісін ќоректік ортаға тўз, гербицид, уыт ќосып, тағы басќа ќолайсыз жағдайда өсіру арќылы төзімділікті арттыруға болады. Гаметоклоналдыќ өзгергіштіктің нәтижесінде гаплоидті өсімдік пайда болуының маңызы зор. Гаплоидті өсімдіктер арасынан мутанттыќ генотипті аныќтап, сўрыптау өте жеңіл. Бидайдың тозаңќап өсіндісінде NaCl тўзына төзімді гаметоклоналдыќ регенеранттар алу мүмкіншілігі зерттеліп тўзға төзімді Ю-580R линиясы алынды. Абсциз ќышќылы енгізілген ќоректік ортасын ќолданып, кўрғаќшылыќќа төзімді АR-45 линиясы сўрыпталды, тағы басќа in vitro әдісінің көмегімен алынған регенерант линиялары Шортандыдағы ғылыми-ґндірістік орталыќта конкурстыќ сорт сынаудан өтуде 20-21-22-23-24-25-суреттер (Бекќожина, 2000, 2005), регенерант-линияларыныѕ RAPD талдау јдісімен ДНК полиморфизмі зерттелуде (26-сурет).
Гаметоклональдыќ ґзгергіштіктіѕ ќауіпті салдары ретінде альбинос ґсімдіктерініѕ пайда болуын айтуєа болады, мысалы бидай, жїгері тозаѕќаптарынан алынєан ґсімдіктен тїссіз ґсімдіктер шыєуы мїмкін. Альбиностар мен олардың жасыл өсімдіктері санының ара салмағы өсімдік түріне, тозаңќаптың даму кезеңіне және өсіру жағдайына байланысты.
Гаплоид организмнің сомалық клет.сыңар хромосомалар жиынтығы.Аталық гаметофитті (тозаңқаптар мен тозаң) ин витро жағдайынан өсіріп,гаплоидтық өсімдіктердіалу андрогенез деп а.
Партеногенез деген аналық жыныс клет.ның ұрықтанусыз көбейіп,организмнің түзілуі. Бірінші рет сасықмендуананың тозаңқаптарын инвитро жағдайында өсіріп,Индияда С.Гуха мен С.Махешвари 1964ж гаплойдтық өсімдіктерді алды.содан кейіносы тәжірибеден француыз ғалымы К.Нич 1967 ж темекінің тозаңкаптарын өсіріп қайталайды. Содан бері гаплойдтардың 200денастам өсімдік түрі алынды.
Билет
In vitro - тірі ағзалардың жасушаларын, ўлпаларын «шыныда», залалсыз жағдайда жасанды ќоректік ортада өсіру әдісі.
2. Микробтық синтез өнімдерін тағамдық мақсаттарда қолдану.
Биообъектілерден, шикізат (субстрат), технологиялық жағдай және құрал– жабдықтардан құралған биотехнологиялық жүйе өндірісте нақты өнім алу үшін тағайындалады. Ашыту, биосинтез және биотрансфорамция процестері нәтижесінде өнім алынады. Биологиялық объектілер арасында ең алдымен микроорганизмдер туралы айту керек, себебі көптеген жұмыс істеп жатқан үлкен маштабты биотехнологиялық өндірістер олардың биопродукциялық активтілігіне негізделген.
Биотехнологиялық жүйеде орталық орынды микроорганизмдер алады, ал технологиялық үрдістің аппараттық жабдықталуы, субстратты таңдауы микробтың биологиялық қасиеттер ерекшеліктеріне, олардың ферментациялауына байланысты. Микробтық клетка эволюцияда биологиялық жүйе ретінде өсіп көбеюге, түрді, популяцияны сақтауға бейімделген. Мицелиалы саңырауқұлақтар және ашытқылар, бактериялармен актиномицеттер қоршаған ортаның биотикалық және абиотикалық факторларымен тұрақты өзара қарым-қатынаста болып келеді, табиғи сұрыпталу әсерінен басқа түрлермен рационалды бейімделуге, симбиозға және бәсекеге түседі, тек қандай да болмасын метоболиттің жоғары өнімдеріне сәйкестенбейді. Сонымен қатар, микроорганизмдер биотехнологиясында ерекше міндеттер басым – адамның тіршілігіне қажетті биологиялық активті қосылыстарды, нақты метоболитті синтездеуге микробтық клетканың табиғи қабілеттілігін жоғарлату, қайта – қайта күшейту болып келеді, тағы ла прокариоттың метоболизм жүйесін қолдана отырып гендік – инженерлік технология жолымен аса құнды эукариоттық ақуыздар алу. Биообъект ретінде микроорганизмдерді айрықша қолдануы келесі критерилерге сүйенеді: микроорганизмдердің жылдам өсіп-көбеюі, биосинтездің көптеген жолдары болуы, жоғары ферметтік белсенділікке ие болғандықтан түрлі органикалық заттарды, ксенобиотиктерді ыдырату қабілеті және керісінше, аэробты және анаэробты жағдайда бейорганикалық заттардан органикалық қосылыстар түзу, микроорганизмдердің барлық жерде кездесуі, экстремалды жағдайларға олардың жылдам бейімделуі (жоғары температура, төменгі рН, осмостық градиентке және т.б.), гендік – инженерлік мутациялар үшін жоғары организмдермен салыстырғанда микробтық клетканың ауқымды технологиялық қол жетерлігі. Қазіргі кезде пробиотиктер мен биологиялық белсенді заттарды түзетін микоорганизмдерді қолдана отырып, қышқыл сүт тағамдарын алудың жаңа түрлерін өндіру биотехнологияның негізгі бағыттарына айналды.Сүт өнеркәсібінің басты міндеттерінің бірі – азық ассортименттерін жетілдіру, ұлттық сүт тағамдарының сапасын арттыру балансалы қоректену талаптарын орындап, халық қолданылатын сүт тағамдарын өндіретін жаңа микроорганизмдер штамдарын өніру және оларды қолданысқа енгізу биотехнологияның басты табтарының бірі болып табылады. Тағамдық өнімдердің технологиясында микроорганизмдер маңызды роль атқарады. Ертедегі биотехнологиялық үрдістің бірі – ол микроорганизмдердің көмегімен ашыту. Ашыту адам өміріне қажетті үрдістерде кеңінен қолданылады және қандайда бір микроорганизмдерді қолдану арқылы іске асырылады. Көбінесе наубайханаларда, шарап, сыра өндірулерде, көкөністерді тұздағанда, сүрлем дайындағанда және сүт өнімдерін сүт қышқылды өнімдерге айналдыру үрдістерінде қолданылады. Ашытудың әр түрін ерекше топтағы микроорганизмдер тудырады. Ашу - ағылшын сөзі «yeast» бұл ескі анғылшын сөзіне «gist», «gyst» «көпіру, қайнау, газ бөлу» деген сөздерінен шыққан. Ашытқылар бұлар ең алғашқы «үй организмдері» болып саналады. Адамдар мың жылдар бой бұларды ферментация мен нан пісіруде қолданған. Археологтар бұрынғы мысыр қалаларынан наубайханалармен нан пісіретін адамдардың бейнелерін тапқан. Археологардың айтуы бойынша мысырлықтар б.з.д 6000 жыл бұрын сыра қайната білген, ал б.з.д 1200 жылы ашыған нан пісіру технологиясын игерген деп есептеледі. Жаңа субстртты ашытуды бастау үшін бұрынғы қалған ашытқыларды қолданған. Осының нәтижесінде әр түрлі шаруашылықта жылдап ашытқы селекциясы жүрді және табиғатта жоқ жаңа физиологиялық рассалар пайда болды. Бұларды ең басында жеке түр деп санаған. Бұлар өсімдік сорттары сияқты, адамның өмір сүру барысында пайда болған өнімдер.
1680 жылы голланд натуралисті Антони ван Левигук ең бірінші болып микроскоп көмегімен ашытқыларды көрді, бірақ оларды ажырата алмады. Тек 1851 жылы француз микробиологы Луи Пастер өзінің «Mémoire sur la fermentation alcoholique» жұмысында сптрттік ашу – бұл химиялық реакция емес (бұрын солай деп ойлаған), бұл ашытқы микроорганизмдердің көмегімен жүретін биологиялық процесс екенін дәлелдеп берді. (1-сурет)
1881 жылы Эмиль Христиан Хансен «Carlsberg»атты дат компаниясының лабораториясының жұмысшысы ашытқының таза культурасын бөліп алды, ал 1883 жылы бірінші рет оны сыра дайындауда қолданған болатын. ΧIΧ ғасырдың соңында Э.Х. Хансен қатысуымен ең бірінші ашытқылар классификациясын жасады. ΧΧ ғасырдың басында ашытқылар коллекциялары мен оны анықтайтын қондырғылар пайда бола бастады. Ал ΧΧ ғасырдың екінші жартысында ашытқылар туралы ғылым (зимоллогия) ашытқының практикалық жағына ғана емес оның экологиясына, цитологиясына және генетикасына көңіл бөле бастады.
ΧΧ ғасырдың отасына дейін ғалымдар тек аскомицетті ашытқылардың жыныстық циклын (шеңберін) бақылаған және бұл барлық сөмкелі саңырауқұлақтардың таксономиялық тобына тән деп есептеген болатын. Жапон ғалымы Исао Банно 1969 жылы Rhodotorula glutinis - тің жыныстық көбею цилын (шеңберін) идуцирлеген болатын, ал бұл базидомицеттерге жатады. Қазіргі кездегі молекулалық-биологияның зерттулері бойынша, ашытқылар аскомицеттер мен базидомицеттер саңырауқұлақтары, бұлар екеуі бір таксонға жатпайды бұлар тек бір өмірлік түрге жатады.
24.04.1996 жылы Saccharomyces cerevisiae - дің ең бірінші эукариотты организмнің геномын (12 млн жүп негіздер) түгелдей секвенирленгені туралы ғалымдар хабарлады. Секвенирлеу 7 жыл уақыт алды және бұған 100 жуық лабораториялар қатысқан болатын. 2002 жылы келесі геномы секвенирленген (13,8 млн жүп негіздерден түратын) ашытқы микроорганизмы және алтыншы эукариотты организм болып Schizosaccharomyces pombe болды.
Сүт қышқылды ашыту адамзатқа ертеден белгілі, алайда оның қандай себептерден, үрдістерден туындайтындығын тек 1857 ж Л.Пастер анықтап дәлелдеген. Қазіргі кезде сүт қышқылды бактериялардың көптеген түрлері белгілі болды. Соның ішінде ен негізгісі кокктар мен ішек таяқшалары.
Сүт қышқылды ашыту микрооганизмдері
Streptococcus lactis. Сүт қышқыл стрептококкасы, барлық сүт тағамдарынан табылған және ол сүттің ұюында басты рөл атқарады. Айран микрофлорасының негізгі құрамдас бөлігі. Бұлар қысқа шынжырша ретінде кейде екі коккалар байланысынан тұрып диплококкалар тәрізді тіршілік етеді. Қолайлы температурададаярланған тағамда (азықта) көбінесе стрептококкалар басым болады. Ал азықтар біраз уақыт сақталса онда таяқшалар көбейеді. Стрептококкалар грам әдісімен боялады, спора жәнекапсула түзбейді, факультативті анаэроб. Сүтте жәнесүт қосылған қорктік ортада жақсы өседі. Ашыту барысында ортада 1%-ке дейін қышқыл түзеді. Бұл микроорганизмнің низин антибиотигін түзетін қасиеті бар. Бұл зат бірқатар микроорганизмдерден, соның ішінде туберкулез ауруын қоздырғыштардың тіршілігін тежей алады.
Lactobacterium bulgaricusm. Бұны алғаш рет болгар айранынан И.И. Мечников бөліп алып зерттеген. Қозғалмайтын таяқша, өсуге қолайлы температура +40-48°С. Ортада 3,0-3,5 %-ке дейін қышқыл түзе алады. Көбінесе ол айран дярлауда қолданылады.
Lactobacterium acidophilum. Бұларды емшектегі балалардың ішегінен бөліп алған. Ол +40°С температурада өсіп - өнеді. Сүтте өскенде 2,2 пайыздай сүт қышқылын тзеді. Көбінесе ацидофилин және ацедофиль айранын даярланғанда қолданылады. Бұлардың шіріту бактерияларын қыра алатын қабілетін зерттеп білген И.И Мечников оларды түрлі ішек – қарын ауруларын емдеуге қолдануды ұсынады.
Lactobacterium delbruckii. Жеке-жеке немесе тізбектеле орналасқан таяқшалар. Спора түзбейді. Тіршілігіне ең қолайлы температура +45°С. Ортада 2,5 % қышқыл түзеді. Ал ортаға ептеп бор салып отырса қышқыл мөлшерін 10 %-ке дейін жеткізуге болады. Бор пайда болған қышқылды бейтраптап, ортадағы қантты бактериялардың толық ашытып шығуына қолайлы жағдай жасайды. Бұл бактериялар басқаларына қарағанда қышқылды көбірек түзетіндіктен сүт қышқылын алу өндірістерінде қолданылып отыр.
Streptococcus cremoris. Кілегей стрептококкасы, торшалары шынжырша орналасқан, сүтте тіршілік етеді, оны ұйытқанда сыртқы түрі қоймалжың қаймаққа тым ұқсас болып келеді. Қаймақ, май, түрлі сырларды (ірімшік) даярлауда ашытқы ретінде қолданылады.
Сыра ашытқысы
Saccharomyces cerevisiae. Сыра ашытқысы немесе жай тағам ашытқысы (лат. Saccharomyces cerevisiae) – бұл бір жасушалы микроскопиялық (диаметрі 5 – 10 микрон) саңырауқұлақ және сахаромицеттер түріне жатады. Бұларды ғылыми зерттеу жұмыстарында қолданылады және олары көбірек қолданылатын жері, тағам өнеркәсібінде алкогольді сусындар (сыра, шарап және т.б.) мен нан пісіру өндірісінде ашытқы ретінде қарқынды қолданады.
Saccharomyces cerevisiae жасушалары вегатативті жолмен бүршіктену және споралар түзу арқылы көбейеді. Ең алғашқыда аналық жасушада төмпешік пайда болады, одан кейін ядро бөлінеді де кейін жасуша қабығы пайда болып жасушалар бір – бірінен ажыратылады. Аналық жасушада бүршіктенуден кейін сызықтар қалады, осы сызықтар арқылы оның жасын анықтауға болады. Қалыпты жағдайда аналық жасуша 20 – 30 бүршік түзе алады. Ашытқы экстрактысын өсіру үшін ең оптималды температура болып 30°С саналады және қоректік ортада пептон мен глюкоза болуы қажет. Осы кезде ашытқы жақсы өсіп - өнеді.
Сыра ашытқылары – бұлар ең жақсы зерттелген организмдердің бірі, осыларды мысал ретінде ала отырып эукариоттық жасушаларды зерттейді. Бұлар тез өсіп - өнеді жәнеадам организмне патогені емес. Ішек таяқшасымен (Escherichia coli) салыстырғанда ашытқы жасушасындағы ДНҚ саны бірнеше рет көп болады және ол күрделірек құрастырылған. Ашытқы жасушаларының өмір сүру қабілеті жоғары тіпті де олардың өзінің генотипінде генетикалық маркер болсада жоғары болады.
Микробтық ақуызды өндіру.
Бәрімізге белгілі, ересек адамға күнделікті дене еңбегінің жүктемесі үшін тағаммен бірге 12,5 кДж (3000 калория) алуы қажет. Энергиядағы бұл қажеттілікті 75 г қант алмастыра алады. Бірақ тамақ бізді тек қана калориямен қамтамасыздандырмайды, сол себепті организмге өсуге және қартайған (солған) жасушалар мен ұлпалардың регенерациясы үшін материалдар қажет, сондықтан тағамның құрамында ақуыздар, майлар, көмірсулар, витаминдер көп мөлшерде болғаны жөн. Адамдар көбінесе жер, мал, балық шаруашылықтарының өнімдерін пайдаланады.
Микроорганизм клеткалары барлық қажетті амин қышқылдарын, оның ішінде алмастырылмайтындарын органикалық және бейорганикалық қосылыстарын синтездеуге қаблетті. Алмастырылмайтын амин қышқылдарының микробты ақуыздағы сандық құрамы жануар және өсімдік текті клеткаларына жақын. Микроорганизм клеткаларында, өсімдік және жануарлардағы сияқты, ақуыздан басқа тағамдық құндылығы бар компоненттер, оның ішінде нуклеин қышқылдары, көмірсулар, амин қышқылдары, органикалық қышқылдар, фосфолипидтер, витаминдер, микроэлементтер және т.б. болады.
Тамақ қосындысы ретінде микроб ақуыздарын қолданудың басты негіздері:
- микроорганизм ақуыздары аминқышқылдық құрамы бойынша адам ақуыздарымен сәйкес келеді;
- құрамында көмір су, липид және витаминдер көп болуы;
- қоректік өсімдіктермен салыстырғанда құрамында амин қышқылдарының көп болуы;
- микроорганизмдердің жоғарғы қарқынмен өсуі мен көбеюі, ақуыз синтезінің жоғары қарқындылығы;
- микроорганизмдер биомассасын өсіру үшін субстрат – бұл ет, сүт, кондитерлік және өндірістің басқа салаларының қалдықтары, оларға оңай қол тиеді және арзан;
- микроорганизмдерді дақылдандыру технологиясы қарапайым, көп сатылы емес, қолайлы жағдайда (30-450 С, рН 3-6), жыл бойына ыңғайлы жағдайда жүреді;
- селекция, мутагенез, рекомбинатты ДНҚ технологиясы жолымен жоғары деңгейлі өнімді штамдар дайындалады.
Тамақ және қоректік ақуыз ретінде ашытқылар (Saccharomyces, Candida), бактериялар (Methylophilus, Pseudomonas, Methylomonas, Acinetobacter), бір клеткалы балдырлар(Spirulina, Chlorella, Scenedesmus) және саңырауқұлақтар (Fusarium) қолданылады.
Ашытқы клеткалар. Saccharomyces cerevisiae, Sacch. carlsbergensis клеткалар құрамында 50 % жуық ақуыз, алмастырылатын және алмастырылмайтын амин қышқылдары, түрлі витаминдер бар; лизин, треонин, валин және лейцин өсімдіктерден асады, бірақ метионин, цистеин аз мөлшерде. Ашытқы биомассасы тиімді субстратта өндіріледі, ал ферментациялау ақуыз өнімін алуда экономды және ұтымды. Saccharomyces cerevisiae ашытқы биомассасынан алынған азықтар алмаспайтын аминқышқылдарының жалпы мөлшері және қатынасы бойынша жоғары қоректік тағамдық өнімдерге көрсетілген талаптарға сай келеді; алмастырылатын және алмастырылмайтын аминқышқылдарының оңтайлы қатынасына ие. Ашытқы клеткалар азот көзі ретінде аммоний тұздарын және аздап жиі нитраттарды қабылдай алады. Биомассаны тез өсіру үшін көміртек көзі ретінде түрлі органикалық субстраттарды пайдалану, бейорганикалық азотты ақуызға айналдыру өнеркәсіптік өндірістің негізі болып табылады.
Ашытқыларды құрамында целлюлозалы шикізаты бар қалдықтардың гидролизаттарында өсіреді. Гидролиз барысында шикізат көзі ретінде ағаш өңдейтін өндірістің қалдықтары, сабан, арпа, мақта қауыздары, жүгері собықтары, қызылша мелассасы, кондитерлік және сүт өнеркәсібінің қалдықтары қолданылады. Гидролиз нәтижесінде микроорганизмдер үшін оңай қорытылатын көмірсу түрлері пайда болады. Ұнтақталған өсімдік шикізаты жоғары атмосфералық қысымда және температурада қышқылды гидролизге ұшырайды. Целлюлоза олиго- және моносахаридтерге дейін гидролизденеді. Гидролизаттан лигнинді бөліп тастайды, минералды тұздар, витаминдер және басқа ашытқы дақылдауына қажетті заттар қосылады.
3. Фермент 1946 жылы американдық химиктер Самнер мен Нортон таза күйіндегі (кристалл) уреаза – ферментін алғаш рет бөліп алғандары үшін Нобель сыйлығын алды. 1963 жылы жануартекті фермент – лизоцимнің біріншілік құрылымы анықталды. Сондай-ақ, ферменттер биологиялық катализатор ретінде ақуыздар екені дәлелденді. Ферменттер молекулярлық массасы 10 мыңнан миллион дальтонға дейінгі ақуыздар болып табылады. Ферменттің атауы - ол әсер ететін затқа байланысты қойылады және –аза- жалғанады. Мысалы, целлюлаза - целлюлоза гидролизін катализдейді, уреаза – мочевина гидролизі және т.б.
Ферменттердің классификациясы олармен катализденетін реакциялардың табиғатына байланысты құралады және 6 кластан тұрады:
1. Оксидоредуктазалар – тотығу-тотықсыздану реакцияларын катализдейтін ферменттер;
2. Трансферазалар – химиялық топтардың тасымалдану реакциясын катализдейтін ферменттер;
3. Гидролазалар – гидролиз реакциясын катализдейтін ферменттер;
4. Лиазалар – химиялық топтардың қосылуын немесе айырылуын катализдейтін ферменттер;
5. Изомеразалар – изомеризация реакцияларын (радикалдардың, атомдардың молекула ішілік орналасуын және т.б) катализдейтін ферменттер;
6. Лигазалар (синтетазалар) – қарапайымдылардан күрделі оргникалық қосылыстарының синтезін катализдейтін ферменттер.
Әдетте метаболизмге қатысатын ферменттер организм жасушасының құрамында болады, сондықтан жасушаішілік ферменттер эндоферменттер деп аталады. Кейбір ферменттер микроорганизм жасушаларымен сыртқы ортаға бөлінеді және жасушадан тыс экзоферменттер деп аталады. Сыртқы ортаға үлкен молекулярлық массасы бар ыдырайтын қосылыстар гидролитикалық ферменттер бөлінеді, олар микроорганизм жасушасына өте алмайды. Әдетте, биотехнологтар жасушадан тыс ферменттерді қолданғанды жөн көреді. Олар өндірістік препараттарға айналуна оңай ұшырайды, өйткені микробты жасушалардың қабырғаларын бұзуды қажет етпейді.
Жануар текті ферменттерді интенсивті биохимиялық үрдістер жүретін органдардан бөледі. Ірі қара мал мен шошқа асқазанының кілегейлі қабаттарынан - пепсин алынады. Шошқаның асқазан асты безінен трипсин, химотрипсин алынады. Бұзау асқазанынан ірімшік өндіруде қолданылатын сычуж ферменттін бөліп алады. Медицина мен биохимия қажеттілігі үшін ферменттерді бұлшық ет, жүрек, талақ, бауыр, тік ішектен (гиалуронидаза ферментін бұқа ендерінен, каталазаны бауырдан, фумаразаны жүректен) бөліп алады. Ірі ет комбинаттарында бауыздалған малдардың органдарынан биохимиялық препараттарды алуға арналған арнайы цехтары бар.
Өсімдік текті ферменттерден амилазамен папаин қолданылады. Папаин - өсімдік протеазасы – қауын ағашының жемісінің құрамында болады. Тек АҚШ-та жыл сайын 100 тоннаға жуық папаин етті жұмсарту үшін қолданылады. Папаин етпен байланысқан кезде бөлме температурасында 2 сағат аралығында байланыс ұлпаларын – коллаген және эластинді ерітеді. Амилаза ферментінің көзі сұлы болып табылады. Сондай-ақ, өсімдік шикізатынан фосфатаза, пероксидаза, уреаза, гемицеллюлаза және т.б алады. Ферментті препараттартың ұлғаюына байланысты өсімдік және жануар шикізаты өндірушілерді қанағаттандырмайды. Өсімдік құрамында ферменттер аз мөлшерде болады. Сонымен қатар, өсімдіктерден ферменттерді алу маусымдық сипатқа байланысты. Жануар органдарын тек ет комбинаттарынан алады, бұл кезде осы шикізатты консервлеумен сақтау жөніндегі проблеммалары туады.
Микробиологиялық жолмен ферменттерді алу мүлдем бөлек жағдай. Микроорганимдер клеткалары сыртқы ортаға бір қатар ферменттерді бөліп шығарады, олар бөлінуі және тазалығынмен айтарлықтай белгіленеді, бұл технологиялық маңызды. Ферменттер макромолекулалар ретінде де белгіленгендіктен, олардың белсенділігі біріншілік құрылысына тәуелді, яғни аминқышқылдарының реттілігінен ірі масштабты химиялық синтез кей жағдайларда қажеттсіз және мүмкін емес. Микроорганизмдер –ферменттерді алудың өте ыңғайлы көзі, өйткені микробтық жасушадағы ферменттер концентрациясы олардың өсу жағдайы мен генетикалық манипуляциясына әсер ету нәтижесінде айтарлықтай жоғарылануы мүмкін. Микроорганизмдердің басқа артықшылығынан өсу жылдамдығы мен қымбат емес қоректік орталарда өсе алатынын, т.б. атап айтқан жөн. Микробтық ферменттер өсімдік пен жануар ферменттерін белсенді түрде алмастырып отыр. Осылай, Bacillus амилазалары сыра өндіру және наубайханалардағы бидай мен сұлы солодына аналогты ферменттерін алмастырды; Aspergillus протеазалар тері дубленуі (жұмсарту) үрдісінде панкреатикалық протеазаларды алмастырды; Mucor – дан алынған ренниндер ірімшік пісіруде т.б. қолданылатын бұзау асқазанының сычуж ферментін алмастырады.
Микроорганизмдер арасында ферменттердің интенсивті синтезделуіне Aspergillus, Trichoderma текті зең саңырауқұлақтары, сондай-ақ Bacillus текті бактериялардың өкілдері және актиномицеттер қабілетті. Микробиологиялық синтезбен аздаған көлемде төрт типті ферменттер өндіріледі: амилазалар, протеазалар, пектиназалар, глюкоизомеразалар. Амилазалар нан пісіруде, спирттік өндірісте (крахмал мен гликоген гидролизі); протеазалар – терілерді жұмсарту үшін, парфюмерияда (тіс пастасы, кремдерге, лосьондарға қоспалар дайындау үшін), синтетикалық жуғыш заттардағы қоспалар ретінде, сондай-ақ медицинада қабыну үрдістерін, күйіктерді, тромбоздарды емдеу үшін; пектиназаларды – пектинді заттардың (полисахаридтердің) созылмалдылығын төмендету үшін, шарап өндіруде – шараптарды мөлдірлендіру, жеміс шырындарын консервілеу кезінде олардың тұнықсыздығын жою үшін; глюкоизомеразалар – глюкозаны фруктозаға изомеризациялау үшін (кока-кола секілді алкоголсыз шарап өндірісінде) қолданылады.
Жасушалармен синтезделетін ферменттердің құрамы мен мөлшері ең алдымен микроорганиздердің тұқым қуалаушылық қасиетіне байланысты, өйткені жасушадағы қалыптасқан әр ақуыздың құрылымы сәйкес генмен анықталады. Ферменттер биосинтезінде генетикалық фактордың анықтаушы рөліне қарамастан, әр фермент бойынша бар технологиялық үрдістердің өндіргіштігі қоректік орта құрамына кіретін көміртегі азот, фосфор және басқа элементтердің көздерінен басқа белгілі фермент биосинтезінің индуктор немесе репрессор рөлін атқаратын заттарға да тәуелді. Солай, липаза ферменті Asp. awamori саңырауқұлағымен синтезделінбейді бірақ қоректік ортаға қоспа ретінде кашалот майын қосқан кезде фермент биосинтезін 100 есе күшейтті. Фермент продуценттерін культивирлеу екі әдіспен жүзеге асырылуы мүмкін: 1) тереңдік әдіс (сұйық қоректік ортада) және 2) беткейлік әдіс (тығыз қоректік ортада).
Культивирлеудің тереңдік әдісі қатаң асептикалық жағдайларда жүргізеді, өйткені асептикалық жағдайдың аз да болсын бұзылуы, фермент түзілуінің толық тоқтатылуына әкеп соғады. Продуцент ферменттерді культивирлеу үшін қоректік орта ретінде көп жағдайларда крахмалдың әр түрлі сорттары (картоп, жүгері, сұлы, күріш), жүгері экстрактысы, соя ұны, ашытқы гидролизаттары және т.б. қолданылады. Себу материалының түрі продуцентке байланысты: саңырауқұлақтар мен актинмицеттер үшін – мицелиалды вегетативті масса, бактериялар үшін – жас өсіп жатқан өсінді. Себінді материалдардың көлемі продуценттің физиологиялық қасиеттеріне байланысты және өндірістік фермент көлемінен 10%-ын құрайды. Өндірістік культивирлеу жұмысына себінді материалдың жасы айтарлықтай әсер етеді. Өте жас себінді өсіндісін қолданған кезде үрдіс көп уақыт алады, ал ескі культура субстраттың рационалды емес қолданысына әкеп соғады. Продуцентті культивирлеу кезінде екі бір-бірімен байланысты үрдіс жүреді – микроорганизм биомассасының ұлғаюы және ортада ферменттердің жиналуы. Фермент биосинтезі ауаның үзіліссіз берілуі мен араластыруы кезінде 2-4 тәулікте өтеді. Егер ферментер экстрацеллюларлы өнімдер болып және сұйық қоректік ортаға бөлінсе онда бұл жағдайда фермент препараттарын биомассаны ажыратқаннан кейін бөліп алады. Кейде ферменттер сыртқы ортаға бөлінбейді және жасушалық органелаллалармен байланыса отыра, жасушадан тек олардың бұзылуы мен сәйкес өңдеуінен кейін ғана бөлініп алынады. Фермент биосинтезі кезінде ортаның оптималды рН көрсеткіші -7.2-7.5, алайда ферменттердің өз оптималды рН әсері әркелкі (рН 2.0-ден –рН 11 дейін). Фермент продуценттердің көбі мезофилді микроорганизмдер болып табылады және олардың даму оптимумы 25-35 С температурада болады. Әдетте, культивирлеу 100 м 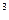 көлемге дейінгі ферментерде жүргізіледі. Ферментерлердің эффектілі жұмысының маңызды жағдайы бақылау және үрдістің негізгі параметрлерінің оптималды деңгейін автоматты ұстап тұру болып табылады.
көлемге дейінгі ферментерде жүргізіледі. Ферментерлердің эффектілі жұмысының маңызды жағдайы бақылау және үрдістің негізгі параметрлерінің оптималды деңгейін автоматты ұстап тұру болып табылады.
Беткейлік культивирлеу әдісі кезінде өсінді тығыз қоректік орта беткейінде өседі. Зең саңырауқұлақтарының мицелилері тығыз бөлшектерді толығымен жабады, жасушалар қоректік заттарды тығыз бөлшек құрамындағы заттар арқылы алады және тыныс алу үшін оттегіні ауадан алады, сондықтан оларды оттегімен қалыпты қамтамасыз ету үшін құрылымы бойынша борпылдақ, қабат биіктігі өте төмен қоректік орталарды қолданады. Мысалы, бидай жармасындағы бір тонна зең саңырауқұлақтарының беткейлік өсіндісін өндіріп шығаратын зауыттар үшін екі тәуліктік өсу кезінде жұқа қабатпен саңырауқұлақ конидияларымен себілген қоректік орта төселінген бөлек 1000-ға жуық кюветтер қажет. Беткейлік әдістің артықшылығы – қоректік орта массасының бірлігіне ферменттің жоғары ақтық концентрациясы (5 кг зең саңырауқұлақтарының беткейлік өсіндісі 100 кг өсінді сұйықтығына жуықтай сәйкес келеді). Беткейлік өсінділер жеңіл бөлінеді және тазартылады. Тығыз қоректік орта негізі қажетті қоректік заттар көзі ретінде бидай жармасы болып табылады. Ылғалдандырылған күйдежарма қоректік ортаның керекті құрылымын қалыптастырады. Кейде жармаға көмірсуға бай материал ретінде қызылшалы жомды, соя шроттарын, соя ұнын (құрамында көп ақуызы бар) қосады. Беткейлік өсіндіні өсіру үрдісі 36-40 сағатқа созылады және 3 периодтан тұрады 1) алғашқы 10-12 сағатта споралардың өсіп-өнуі; 2) келесі 12-14 сағатта ақ-сұр түсті үлпілдеген налет ретінде мицелилердің өсуі; 3) соңғы 12-18 сағатта – ферментация. Технология бойынша өсінділерді өсіру кюветтерде жүреді. Олар перфорирленген түбімен мырышталған темірден жасалады. Олардың көлемі әдетте 2х20 мм саңылау түрінде перфорациясы бар 600х900х30 мм болады. Толтырылған кюветтерді көпярусты жылжымалы камераларға (бір камераға 700 кг қоректік орта сияды) орналастырады. Өскен өсінді өсіп кеткен мицелилермен тығыз байланысқан үлбіреген бөлшектерден құралған пласт ретінде болады. Бұл массаны арнайы дезинтеграторларда 5-6 мм көлемдегі гранулаларға дейін ұсақтайды. Ылғалдылығы 40-50% болатын ұсақталған өсіндініарнайы конвективті кептіргіштерде 10-12%-ға дейін кептіреді.
Беткейлік әдіспен өсірілген микроорганизмдер культурасын және тереңдік культивирлеуден кейінгі өсінді сұйықтықтардың құрамында көп көлемде балласт заттар болады (продуцент биомассасы, қолданылмаған қоректік орта компоненттері, метаболизм өнімі). Ферменттің өзіндік үлесі беткейлік үшін -1% және тереңдік өсінділер үшін -0.1% құрайды. Фермеенттерді бөліп алу және тазарту қиын және қымбат үрдіс. Сондықтан кейбір жағдайларда ферментті препараттарды тазартылмаған күйде қолданады (тері-тамырлы өндірісте). Сол уақытта тағам өндірісінде, медицинада тек жоғары деңгейде тазартылған ферменттер қолданылуы мүмкін. Ферментті препараттардың стандартизациясы – препараттың белсенділігін әр түрлі текті толтырғыштарды (крахмал, лактоза) қолданумен ГОСТ-пен анықталатын стандартқа дейін жеткізу соңғы кезең болып табылады.
Иммобилизирленген ферменттер. Ферменттерді адамдардың әр түрлі практикалық салаларында дәстүрлі қолданады: тағам, фармацевтикалық, тоқыма (текстиль), терітамыр және т.б. өндіріс салаларында, медицинада, ауыл шаруашылығында, органикалық синтезде, химиялық анализде және т.б. Сонымен қатар, қолданбалы энзимология дамуына көп уақыт бойы қымбаттылығымен немесе әлем нарығында керекті ферменттердің әсіресе олардың таза препараттардың мүлдем болмауы кедергі жасап тұрды. Ферменттердің кең қолдану мүмкіндігі екі себепке байланысты қиындатылды. Біріншіден, ферменттер сақтау барысында тұрақсыз, сонымен қатар әр түрлі әсерлер кезінде, әсіресе жылулық. Екіншіден, ферменттердің реагенттер мен өнім реакцияларынан қиын бөліп алынуына көп қолданылуы қиындатылған болатын. Қолданбалы энзимологияда жаңа адымдар иммобилизденген ферменттерді құру нәтижесінде қалыптастырылды. Иммобилизация – фермент молекуласы қозғалғыштығының, олардың конформационды қайта құрастыруларының оқшаулануы – оның молекуласының белсенді орталығын өзінің жұмыс қабілетін (каталитикалық белсенділігі) ұзақ уақыт бойы, құрылымды өзгертілулерге және конфигурациясына әкелмейтін, фермент құрылысын бекітуге жағдай жасау үшін физика-химиялық принциптерге негізделген.
Иммобилизденген ферментті препараттар өздерінің натифті ізашарларымен салыстырғанда айтарлықтар бір қатар артықшылықтары бар: біріншіден, катализаторларды реакцияларды орталардан жеңіл бөліп алуға болады, бұл мүмкіндік береді: а) реакцияны керекті сәтте тоқтату; б) катализаторды қайта қолдану; в) ферментпен ластанбаған өнім алу. Екіншіден, катализаторларды қолдану ферменттерді үздіксіз жүргізуге жағдай жасайды, мысалы, ағынды колоналарда, және катализденіп жатқан реакцияны жылдамдығын реттеу, сондай-ақ ағын жылдамдығының өзгеруі арқылы өнімнің шығуы.
Иммобилизденген ферменттерді алу үшін органикалық және бейорганикалық тасығыштардың көбін қолданады. Материалдарға қойылатын негізгі талаптар келесідей: 1) жоғары химиялық және биологиялық тұрақтылық; 2) жоғары механикалық беріктігі; 3) фермент үшін қажеттілікті өтімділік; 4) гранула, мембрана және т.б. түрінде алу мүмкіндігі; 5) жоғары гидрафильділігі; 6) төмен құны.
Көп жағдайда иммобилизация үшін полисахаридті органикалық тасығыштарды қолданады – целлюлоза, декстран, агароза және оның туындылары. Целлюлозаның негізгі өндірістік маркалары: аминоэтилцеллюлоза, фосфорилцеллюлоза, диэтиламинцеллюлоза, бромацетилцеллюлоза, ДЭАЭ-целлюлоза. Декстрандар – бактериалды көздерден алынған полисахаридтер, «сефадекс» атымен шығарылады. Әр түрлі сефадекстер тігілім дәрежесі және, сәйкесінше, үлбіреу және саңылаулығы бойынша ажыратылады. «Сефароза» атты агароза препараттары өндіріледі. Ақуызды тасығыштардың (каллоген, желатин, кератин) негізгі қасиеттері ферменттерге қатысы бойынша жоғары сыйымдылығы болып табылады, сондай-ақ және олардың көбін (фибриллярлы табиғаты арқасында) жұқа пленка (мембрана жуандығы 80 мкм) ретінде қолданылуы. Коллаген – фибриллярлы ақуыз, сіңірлер мен шеміршектердің негізгі компоненті, бөлінудің жоғары тұрақтылығына ие. Желатин – қайта өңделген коллаген өнімі – токсиндік еместігі мен жеңіл биодеградация әсерінен фармакология мен тағам өндірісінде кең қолданады. Табиғи липидтерден иммобилизациялау үшін биомембрана компонентерің қолданады – лецитиндер, кардиолипид, сфингомиелин. әдетте липидті тасығыштарды әр түрлі беткейлерде (су, силикагель, ыс) моноқабат немесе биоқабат (сфералық формалар) – липисомалар ретінде қолданылады.
Қол жетімді синтетикалық палимерлерден кең түрлілігі олардың ферменттердің иммобилизациясы үшін тасығыш ретінде қолданылуын қамтамасыз етеді. Полимерлі молекуларға әр түрлі функционалды топтарды енгізген кезде кең көлемде тасығыштын физикалық қабілеттерін өзгеруге болады. Бейорганикалық тасығыштардан кең қолданылатыны: силикогель, саз балшық, керамика, табиғи минералдар, графиттирленген ыс (сажа), металдар және олардың окситтері.
Иммобилизация әдістері: адсорбция және химиялық таңбалау (ковалентті байланысқан) жолымен иммобилизациялау; полимерлі құрылысқа қосу жолымен иммобилизациялау; инкапсулирлеу жолымен иммобилизациялау: кесе көлденең таңбалау жолымен иммобилизациялау.
Қазіргі уақытта иммобилизденген ферменттердің қатысуымен жүргізілетін жеті үрдіс тағам өндірісінде кең ауқымды қолданыс тапты: 1) глюкоза-фруктозды сироптарды және глюкозадан фруктозаны өңдіру; 2) оптикалық белсенді L-амин қышқылдардың өздердің рацемикалық қоспаларынан алу; 3) фумар қышқылынан аспаргин қышқылының синтезі; 4) фумар қышқылынан алма қышқылының синтезі; 5) диеталық лактозасыз сүт өңдірісі; 6) сүт сарысуынан қанттарды алу; 7) пенициллин қатарының жартылай синтетикалық антибиотиктерінің өндірісі үшін қарапайым пенициллиннен 6-аминопеницилланды қышқылды (пенициллинді ядро) алу.
Ферменттерді иммунохимиялық әдіс анализінің сезімталдылығын жоғарлату үшін қолданған кезде қызықты мүмкіндіктер анықталды. Антиген-антидене кешенің анықтау үшін иммунохимиялық реакцияларда әдетте жеңіл детектирленетін таңбаны қолданады – радиоактивті атом, ол компоненттердің біреуіне (радиоактивті йод, тритий) енгізіледі. Радиоактивті таңбалаудың әдісінің сезімталдылығы бойынша оның каталитикалық белсенділігі анықталады. Бұл әдісті қолдану бойынша жеткілікті материал жиналды, ол иммуноферментті талдау (ИФТ) деген атқа иеленді. ИФТ көмегімен антиген қабілетіне ие әр бір заттар, соның ішіне көптеген адам, жануар және өсімдік ауа қоздырғыштары детектирленуі мүмкін.








