 2015-04-06
2015-04-06 1020
1020Традиционно у лишайников различают три способа размножения: половое, бесполое
и вегетативное.
В результате полового процесса у лишайников формируются плодовые тела:
апотеции, перитеции, гастеротеции. Лишайник развивается только в том случае, если спора
на месте прорастания встречает нужную водоросль.
При определении лишайников форма плодового тела, цвет, строение являются
важными таксономическими признаками.
Самый простой способ - перенос обломков слоевища ветром, водой или животными
на другие места. Особыми приспособлениями для размножения лишайников являются
соредии и изидии.
Бесполое размножение. Б р происходить с помощи пенидии. Пенидии - это вместилище внутри таллома которое развиваются конидии (грибная часть).
Вегетативное размножение
Происходить соредиями (соредия –мелкие частицы образ-ые одними или несколькими клетками водорослей оплетенные грибами). Соредии формируется сораля, образ виде выростов. Сорали – сов-ть соредий. в якутии насчитывается 550 видов. Ведущие сем-ва Cladoniceae, Parmeliaceae.
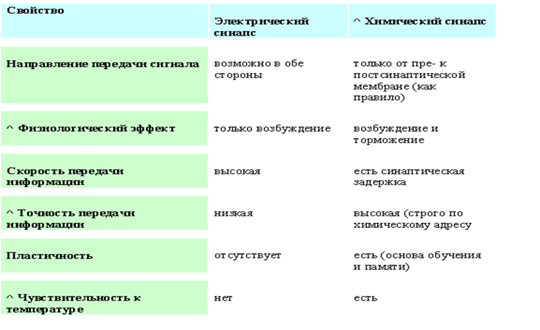
Основные структурно-функциональные элементы нейрона, тело нейрона, дендриты, аксон. Типы нейронов. Механизмы связи между нейронами. Электрический и химический синапсы, их принципиальные отличия.
Структурной и функциональной единицей нервной системы является нервная клетка — нейрон.
Нейроны — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов. Уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний — синапсов.
Выполнению функций нейрона способствует синтез в его аксоплазме веществ-передатчиков — нейромедиаторов (нейротрансмиттеры): ацетилхолина, катехоламинов и др. Размеры нейронов колеблются от 6 до 120 мкм.
Число нейронов мозга человека приближается к 10 в 11 степени. На одном нейроне может быть до 10 000 синапсов. Если только эти элементы считать ячейками хранения информации, то можно прийти к выводу, что нервная система может хранить 10 в 19 степени ед. информации, т. е. способна вместить практически все знания, накопленные человечеством.
Строение нейрона. Функционально в нейроне выделяют следующие части: воспринимающую — дендриты, мембрана сомы нейрона; интегративную — сома с аксонным холмиком; передающую — аксонный холмик с аксоном.
Тело нейрона (сома), помимо информационной, выполняет трофическую функцию относительно своих отростков и их синапсов. Перерезка аксона или дендрита ведет к гибели отростков, лежащих дистальней перерезки, а следовательно, и синапсов этих отростков. Сома обеспечивает также рост дендритов и аксона.
Дендриты — основное воспринимающее поле нейрона. Мембрана дендрита и синаптической части тела клетки способна реагировать на медиаторы, выделяемые аксонными окончаниями изменением электрического потенциала.
Обычно нейрон имеет несколько ветвящихся дендритов. Необходимость такого ветвления обусловлена тем, что нейрон как ин- формационная структура должен иметь большое количество входов.
Информация к нему поступает от других нейронов через специализированные контакты, так называемые шипики.
«Шипики» имеют сложную структуру и обеспечивают восприятие сигналов нейроном. Чем сложнее функция нервной системы, чем больше разных анализаторов посылают информацию к данной структуре, тем больше «шипиков» на дендритах нейронов. Максимальное количество их содержится на пирамидных нейронах двигательной зоны коры большого мозга и достигает нескольких тысяч. Они занимают до 43% поверхности мембраны сомы и дендритов.
Аксон представляет собой вырост цитоплазмы, приспособленный для проведения информации, собранной дендритами, переработанной в нейроне и переданной аксону через аксонный холмик — место выхода аксона из нейрона.
Аксон данной клетки имеет постоянный диаметр, в большинстве случаев одет в миелиновую оболочку. Аксон имеет разветвленные окончания. В окончаниях находятся митохондрии и секреторные образования.
Типы нейронов Строение нейронов в значительной мере соответствует их функциональному назначению. По строению нейроны делят на три типа: униполярные, биполярные и мультиполярные.
Истинно униполярные нейроны находятся только в мезэнцефалическом ядре тройничного нерва. Эти нейроны обеспечивают проприоцептивную чувствительность жевательных мышц.
Другие униполярные нейроны называют псевдоуниполярными, на самом деле они имеют два отростка (один идет с периферии от рецепторов, другой — в структуры центральной нервной системы). Оба отростка сливаются вблизи тела клетки в единый отросток. Все эти клетки располагаются в сенсорных узлах: спинальных, тройничном и т. д. Они обеспечивают восприятие болевой, температурной, тактильной, проприоцептивной, бароцептивной, вибрационной сигнализации.
Биполярные нейроны имеют один аксон и один дендрит. Нейроны этого типа встречаются в основном в периферических частях зрительной, слуховой и обонятельной систем. Биполярные нейроны дендритом связаны с рецептором, аксоном — с нейроном следующего уровня организации соответствующей сенсорной системы.
Мультиполярные нейроны имеют несколько дендритов и один аксон. В настоящее время насчитывают до 60 различных вариантов строения мультиполярных нейронов, однако все они представляют разновидности веретенообразных, звездчатых, корзинчатых и пирамидных клеток.
Классификация нейронов Существует классификация нейронов, учитывающая химическую структуру выделяемых в окончаниях их аксонов веществ: холинергические, пептидергические, норадреналинергические, дофаминергические, серотонинергические и др.
По чувствительности к действию раздражителей нейроны делят на моно-, би-, полисенсорные.
Моносенсорные нейроны. Располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей сенсорности. Например, значительная часть нейронов первичной зоны зрительной области коры большого мозга реагирует только на световое раздражение сетчатки глаза.
Бисенсорные нейроны. Чаще располагаются во вторичных зонах коры какого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности.
Полисенсорные нейроны. Это чаще всего нейроны ассоциативных зон мозга; они способны реагировать на раздражение слуховой, зрительной, кожной и других рецептивных систем.
Функционально нейроны можно также разделить на три типа: афферентные, интернейроны (вставочные), эфферентные
Афферентные нейроны — нейроны, воспринимающие информацию.
Вставочные нейроны, или интернейроны, обрабатывают информацию, получаемую от афферентных нейронов, и передают ее на другие вставочные или на эфферентные нейроны.
Эфферентные нейроны нервной системы — это нейроны, передающие информацию от нервного центра к исполнительным органам или другим центрам нервной системы.
В отличие от других клеток организма нейроны обладают длинными отростками (аксонами и дендритами), позволяющими им контактировать между собой и с окружающими клетками (у млекопитающих длина аксонов может быть более метра).
Концентрация некоторых ионов (в первую очередь ионов калия, натрия, кальция и хлора) внутри отростков и в окружающей их среде различна. За счет этого появляется разность потенциалов по отношению к мембране аксона. Ионы могут проникать внутрь отростков и выходить из них через специальные белковые ионные каналы, пронизывающие внешние стенки аксонов.
Таким образом, вдоль аксона одного нейрона нервный импульс передается как электрический сигнал: в виде изменения разности потенциалов (сами ионы вдоль отростка не перемещаются, а служат только для создания электрического напряжения).
В начале 1970-х годов было показано, что некоторые нейромедиаторы, такие как серотонин, норадреналин, дофамин, действуют в нервной системе по механизму, совершенно отличному от быстрой синаптической передачи.
Американский биохимик Пол Грингард (Paul Greengard) установил, что эти медиаторы не просто изменяют мембранный потенциал, а на глубоком уровне влияют на обмен веществ в самом нейроне, приводя к длительным изменениям в способности синапсов проводить сигналы.
Это явление было названо непрямой (или медленной) синаптической передачей, ответственной за такие сложные свойства нервной системы, как эмоции и память. Эффекты, связанные с медленной передачей, получили название метаботропные.
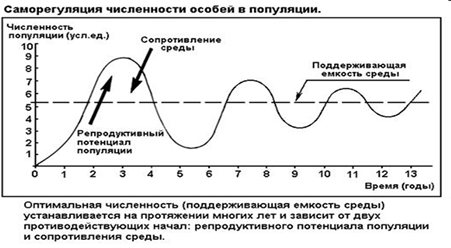
3. Популяция. Статические характеристики: общая численность, плотность, структура - пространственная половая возрастная
Популяция – это часть вида (состоит из особей одного вида), занимающая относительно однородное пространство и способная к саморегулированию и поддержанию определенной численности.
Как первая надорганизменная биологическая система, популяция обладает определенной структурой и свойствами.
Структуру популяции отражают такие ее показатели, как:
численность и распределение особей в пространстве,
соотношение групп по полу и возрасту,
их морфологические, поведенческие и другие особенности.
Численность — общее количество особей в популяции. Эта величина характеризуется широким диапазоном изменчивости, однако она не может быть ниже некоторых пределов. Сокращение численности по сравнению с этими пределами может привести к вымиранию популяции. Полагают", что если численность популяции меньше нескольких сотен особей, то любые случайные причины могут сократить ее настолько, что оставшиеся особи не смогут встречаться и оставить потомство. Рождаемость перестанет покрывать естественную убыль, и оставшиеся особи в течение сравнительно короткого времени вымрут.
Плотность — число особей на единицу площади или объема. При увеличении численности плотность популяции, как правило, возрастает; она остается прежней лишь в случае ее расселения и расширения ареала. У некоторых животных плотность популяции регулируется сложными поведенческими и физиологическими механизмами.
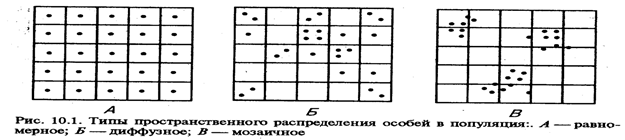
Пространственная структура популяции характеризуется особенностями размещения особей на занимаемой территории. Она определяется свойствами местообитания и биологическими особенностями вида. Наряду со случайным и равномерным распределением в природе наиболее часто встречается групповое распределение. Группа животных, прилагая совместные усилия, может легче защищаться от хищников, искать и добывать корм. Жизнь в семьях, стадах, колониях, гаремах приводит также к групповому распределению особей. Пространственная структура может изменяться во времени; она зависит от сезона года, от численности популяции, возрастной и половой структуры и т. д.
Половая структура отражает определенное соотношение мужских и женских особей в популяции. Генетический механизм определения пола обеспечивает расщепление потомства по полу в соотношении 1: 1.
Изменение половой структуры популяции отражается на ее роли в экосистеме, так как самцы и самки многих видов отличаются друг от друга по характеру питания, ритму жизни, поведению и др. Так, самки некоторых видов комаров, клещей и мошек являются кровососущими, в то время как самцы питаются соком растений или нектаром. Преобладание доли самок над самцами обеспечивает более интенсивный рост популяции.
Возрастная структура отражает соотношение различных возрастных групп в популяциях, зависящее от продолжительности жизни, времени наступления половой зрелости, числа потомков в помете, количества потомств за сезон и др. Если какая-либо возрастная группа сокращается либо увеличивается, это сказывается на общей численности популяции.

2. Побег и его основные функции. Ветвление и нарастание системы побегов. Многообразие внутреннего строения стеблей семенных растений.
Побег – это стебель, несущий листья и почки. Побег имеет метамерное строение. Один метамер состоит из узла, междоузлия, листочка и почки.
Узел – это место прикрепления листа
Междоузлие – участок стебля между двумя узлами.
Стебли и листья имеют единое происхождение как в онтогенезе (из конуса нарастания), так и в филогенезе (из телома).
Функция побега: поддержание жизни.
В онтогенезе побег образуется из зародыша семени
Ветвление играет очень большую роль. Благодаря ему происходит увеличение фотосинтезирующей поверхности, захват территории. У растений может размножение, как вегетативное, так и семенное.
Ветвление происходит за счет боковых почек. Но у некоторых растений сохраняется верхушечное ветвление, которое было у древесных растений. Н-р: у ринии.
У современных растений 2 типа ветвления:
- верхушечное (дихотомическое)
- боковое
При верхушечном ветвлении происходит раздвоение конуса нарастания. В результате образуются 2 конуса нарастания и из каждого конуса нарастания образуются ветви. Эти ветви не являются боковыми, их называют ветвями I порядка, потом ветви II порядка.
Такое ветвление еще называют дихотомическим. Это ветвление сохранилось у плаунов, селагинеллы, некоторых папоротниковых.
У остальных будет боковое ветвление. Боковое ветвление осуществляется за счет боковых почек. При ветвлении увеличивается количество ветвей и образуется система побега. Система побегов нарастает разным способом.
Ветвление – это увеличение количества одноименных структур.
Нарастание – это линейный рост побегов (рост в длину).
Типы нарастания:
- дихотомическое – связано с верхушечным ветвлением. Более древний тип.
- моноподиальное – при этом формируется одна главная ось за счет многолетнего функционирования конуса нарастания. На этой оси образуются боковые почки. Из них развивается боковое ветвление. Верхушечная почка ежегодно дает новые побеги. Моноподиально нарастают голосеменные и многие травянистые растения (подорожник).
- симподиальное – при этом растение не образует одну монолитную главную ось. Главная ось состоит из отдельных симподий или побегов замещения. При симподиальном нарастании побега главная ось очень рано перестает расти, т.к. отмирает верхушечная почка. Вместо верхушечной почки или верхушки стебля занимает боковая почка путем перевершинивания. Более продвинутый тип. Характерно для покрытосеменных растений: береза, ива, кустарники, кустарнички, многолетние травянистые растения.
Симподиальные типы обладают большей выживаемостью. Формирует больше боковых ветвей, больше генеративных побегов.
- ложнодихотомическое – верхушечная почка формирует цветок или соцветие. На этом рост этой оси прекращается. Это ветвление называется ложной, потому что конус нарастания образует цветок.
Анатом стр стеблей 1 и 2 дольных раст. На поперечном срезе стебель можно разделить на неск зон тканей: зона покров тк, первичн коры и центр-го цилиндра. В состав первич коры входят: колленхима, осн тк, эндодерма. Зоне центр цил нах-ся тк: осн тк, склеренхима, провод тк, стебель от корней отлич-ся присут сердцевины.
Стебель двудольных
1.Наличие первичной коры
2.Есть камбий
3.Вторичный рость стебля (в толщину)
4.Проводящие пучки открытые
5.Все пучки располагаются по кругу
Стебель однодольных
1.Первичная кора отсут.
2.Нет камбия
3.Вторичный рост отсут.
4.Проводящий пучок закрытый
5.Пучки располагаются рассеянно.
Для всех стеблей характерно присутствие сердцевины. Для травянистых характерен пучковой тип строения стебля.
Стебель древесных растений: Для всех древесных характерен не пучковый тип строения стебля; образование годичных колец; есть луб (вторичная кора) – в ней есть вторичная флоэма: есть сердцевина.
Голосеменные
1.Ксилема состоит из трахеид
2.Флоэма (дуб) состоит их ситовидных кл.
3.Есть смоляные каналы
4. сердцевинные лучи сост из 2х типов клеток: лучевой паренхимы и лучевых трахеид.
Покрытосеменные
1.Ксилема состоит из трахей и трахеид
2.Флоэма (дуб) состоит ситовидных трубок с кл. спутниками.
3.Отсутст. смоляные каналы
4. сердц-е лучи сост из лучевой паренхимы.
Транспорт веществ в растении. Восходящий и нисходящий токи веществ в растениях. Передвижение органических веществ. Транспортные формы веществ. Зависимость транспорта от температуры, водного режима и минерального питания.
Клетки обмениваются различными веществами с окружающей их средой в результате диффузии. Однако перенос веществ обычной диффузией на большие расстояния неэффективен; возникает необходимость в специализированных системах транспорта. Такой перенос из одного места в другое осуществляется за счёт разности давлений в этих местах. Все переносимые вещества движутся с одинаковой скоростью в отличие от диффузии, где каждое вещество движется со своей скоростью в зависимости от градиента концентрации.
У сосудистых растений передвижение веществ осуществляется по двум системам: ксилеме (вода и минеральные соли) и флоэме (органические вещества). Передвижение веществ по ксилеме направлено от корней к надземным частям растения; по флоэме питательные вещества движутся от листьев.
Одним из важнейших механизмов транспорта веществ в растении является осмос. Осмос – это переход молекул растворителя (например, воды) из областей с более низкой концентрацией в области более с высокой концентрацией через полупроницаемую мембрану. Численно осмос характеризуется осмотическим давлением – давлением, которое нужно приложить, чтобы предотвратить осмотическое поступление воды в раствор.
В растениях роль таких полупроницаемых мембран играют плазматическая мембрана и тонопласт (мембрана, окружающая вакуоль). Если клетка контактирует с гипертоническим раствором, то вода начинает выходить из клетки наружу. Этот процесс называется плазмолизом. Клетка при этом сморщивается. Плазмолиз обратим: если такую клетку поместить в гипотонический раствор (с более высоким содержанием воды), то вода начнёт поступать внутрь, и клетка снова набухнет. При этом внутренние части клетки (протопласт) оказывают давление на клеточную стенку. У растительной клетки набухание останавливается жесткой клеточной стенкой.
Основная масса воды поглощается молодыми зонами корней растений в области корневых волосков – трубчатых выростов эпидермиса. Благодаря им значительно увеличивается всасывающая воду поверхность. Вода поступает в корень за счёт осмоса и движется вверх к ксилеме по апопласту (по клеточным стенкам), симпласту (по цитоплазме и плазмодесмам), а также через вакуоли. Надо заметить, что в клеточных стенках имеются полоски, называемые поясками Каспари. Они состоят из водонепроницаемого суберина и препятствуют продвижению воды и растворённых в ней веществ. В этих местах вода вынуждена проходить через плазматические мембраны клеток; полагают, что таким образом растения защищаются от проникновения токсичных веществ, патогенных грибов и т. п.
Подъём воды по ксилеме происходит, по-видимому, за счёт испарения воды в листьях. В процеcсе испарения в кроне образуется недостаток воды. Поверхностное натяжение в сосудах ксилемы способно тянуть вверх весь столб воды, создавая массовый поток. Скорость подъёма воды составляет около 1 м/ч (до 8 м/ч в высоких деревьях); чтобы поднять воду к вершине высокого дерева, требуется давление порядка 40 атм. Следует иметь в виду, что одни только капиллярные эффекты способны поднять воду на высоту не более 3 м.
Вторая важная сила, участвующая в подъёме воды, – это корневое давление. Оно составляет 1–2 атм (в исключительных случаях – до 8 атм). Этой величины, конечно, недостаточно, чтобы в одиночку обеспечить движение жидкости, но её вклад у многих растений несомненен.
Попадая по ксилеме в листья, вода и минеральные вещества распределяются через разветвлённую сеть проводящих пучков по клеткам. Движение по клеткам листа осуществляется, как и в корне, тремя способами: по апопласту, симпласту и вакуолям. На свои нужды растение использует менее 1 % поглощаемой им воды, остальное в конце концов испаряется через восковый слой на поверхности листьев и стеблей – кутикулу (около 10 % воды) – и особые поры – устьица (90 % воды). Испарение воды (транспирация) осуществляется за счёт энергии солнца.
На скорость испарения влияют многие факторы; как внешние условия (свет, температура, влажность, наличие ветра, доступность воды в почве), так и особенности строения листьев (площадь поверхности листа, толщина кутикулы, количество устьиц). Ряд внешних факторов приводит к уменьшению диффузии воды из листьев, другие (например, отсутствие света или сильный ветер) вызывают замыкание устьиц (благодаря работе особых замыкающих клеток). Растения засушливых регионов имеют специальные приспособления для уменьшения транспирации: погруженные глубоко в листья устьица, густое опушение из волосков или чешуек, толстый восковой налёт, превращение листьев в колючки или иглы и другие. Осенний листопад в умеренных широтах также призван уменьшить испарение воды, когда наступят холода.
Из почвы растение получает не только воду, но и минеральные соли. Эти вещества движутся в корнях под действием диффузии. За счёт энергии дыхания возможен также их активный транспорт против градиента концентрации. Попадая в ксилему, минеральные вещества разносятся по всему организму с массовым током воды.
Некоторые минеральные вещества, выполнив свою полезную функцию, могут перемещаться дальше вверх или вниз по флоэме. Это происходит, например, перед сбрасыванием листьев, когда накопленные листьями полезные вещества сохраняются, откладываясь в других частях растения.
У многоклеточных растений есть ещё одна транспортная система, предназначенная для распределения продуктов фотосинтеза, – флоэма. В отличие от ксилемы, органические вещества могут транспортироваться по флоэме и вверх, и вниз. 90 % переносимых веществ составляет сахароза, которая практически не участвует в метаболизме растения непосредственно и поэтому является идеальным углеводом для транспорта.








