 2015-05-05
2015-05-05 1126
1126Сложность структуры системы определяется числом п ее элементов и числом т связей между ними. Если в какой-либо системе исследуется число частных дискретных состояний, то сложность системы С определяется логарифмом числа связей:
C = lg m. (1)
Системы условно классифицируются по сложности следующим образом:
1) системы, имеющие до тысячи состояний (О <С < 3), относятся к простым,
2) системы, имеющие до миллиона состояний (3 < С < 6), являют собой сложные системы,
3) системы с числом состояний свыше миллиона (С > 6) идентифицируются как очень сложные.
Все реальные природные биосистемы очень сложны. Даже в структуре единичного вируса число биологически значимых молекулярных состояний превышает последнее значение. Кроме того, существует и другой критерий сложности, связанный с поведением системы, ее реакцией на внешнее воздействие. Если система способна к акту решения, т.е. к выбору альтернатив поведения (в том числе и с помощью случайного механизма), то такая решающая система считается сложной. Сложной будет и любая система, включающая в себя в качестве подсистемы хотя бы одну решающую систему.
2. Разнообразие состава или взаимосвязей в системе оценивается показателем Е. Симпсона
Ds = 1 - 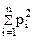
или показателем К. Шеннона
Hs = – 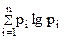
где Ds, Hs — индексы разнообразия;
pi — относительная численность (частота встречаемости) i-й разновидности элементов в совокупности п разновидностей (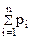 = 1).
= 1).
Эти показатели важны при количественной оценке биоразнообразия экосистем и принципиально отличаются от числа представленных в экосистеме видов.
Например, пусть существуют два одинаковых по суммарной численности (100) набора состава древостоя: А — сосна (35), береза (35), ель (30); Б — сосна (90), береза (3), ель (2), осина (2), пихта (1), дуб (1), клен (1). Рассчитанные в соответствии с показателями Симпсона (2.2) индексы разнообразия равны соответственно DA = 0,665 и DБ = 0,188. Те же индексы, рассчитанные по Шеннону, т.е. по формуле (2.3), соответственно равны HA = 0,548 и HБ = 0,216. Хотя в составе Б вдвое больше видов, но собственно разнообразие в 2,5—3,5 раза меньше, так как это типичный сосняк с малой примесью других видов, тогда как состав А — типичный смешанный лес.
3. Вычисление разнообразия по формуле К. Шеннона — это частный случай расчета количества информации для оценки неопределенности функционирования системы:
H = – 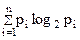
где pi — вероятность реализации i-того события;
-log2 pi — количество информации, соответствующее i-му событию.
Это выражение приводит к интерпретации такого физического понятия, как энтропия, — важного параметра состояния любой динамической системы. В аналогичном вероятностно-информационном определении энтропии S символ pi обозначает вероятность того, что система находится в некотором i-м состоянии:
где kБ - постоянная Больцмана, равная 1,3807 • 10-23 Дж/град.
Энтропия как мера необратимого рассеяния (диссипации) энергии при ее превращениях и вероятность пребывания системы в данном термодинамическом состоянии (принцип Л. Больцмана) выступает здесь и как мера вероятностной неупорядоченности, хао-тизации элементов системы (Волькенштейн, 1984).
4. При условии, что для каждого элемента системы возможны два состояния (основное и возбужденное) и они равновероятны (в этом случае -log2 pi = 1 бит информации), отношение IS = H/S может служить в качестве информационного эквивалента энтропии: IS = 4,38 • 1023 бит на одну энтропийную единицу или 1,045 • 1023 бит наДж/К1.
Поскольку
∆S = ∆Q/T,
где Q — количество теплоты (энергии);
Т — абсолютная температура, знание которой позволяет оценить информационный эквивалент энергии:
IЕ = 1,045 • 1023 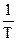 бит/Дж.
бит/Дж.
Это означает, что в энергетическом отношении структурная информация сама по себе чрезвычайно «дешева». Ее обратная зависимость от температуры имеет скрытое универсальное значение и представляет самостоятельный интерес.
Сама по себе энтропия и тем самым неопределенность состояния системы может только возрастать, т.е. информация сама по себе может только утрачиваться. Только в открытых системах (с накачкой), отдающих энтропию, информация может приобретаться. Поэтому переработку информации можно рассматривать как некую частную разновидность самоорганизации.
5. Оценка относительной организации системы R, зависящая от сложности и разнообразия состава, рассчитывается по формуле:
R= 1 – 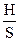
По этому параметру системы также разделяются на три группы:
1) если оценка R мала (0 < R < 0,1), то система считается вероятностной, неустойчивой, обладающей малой жесткостью и способной гибко изменять свои состояния;
2) если значение R сравнительно велико (0,3 < R <1), то система считается детерминированной, т.е. консервативной, жесткой, устойчивой;
3) промежуточное положение занимают квазидетерминированные системы (0,1 <R < 0,3).
Большинство природных биосистем имеет вероятностный или квазидетерминированный характер.
Организмы обладают иерархией структур и функций, в которой по мере усложнения организации возрастает разнообразие системных качеств. В ходе их эволюции образуются и приобретают все большее значение выделенные структуры управления. В отличие от них природные вероятностные системы, состоящие из большого числа слабо скоррелированных индивидуумов (экосистемы), могут обладать функциональной иерархией особей, но не нуждаются в выделенной внутренней системе управления. Они способны к самоподдержанию и во многих случаях без каких бы то ни было «центральных регуляторов» обнаруживают удивительно тонкую и точную авторегуляцию.

