2015-05-06
2015-05-06 13287
13287Процесс развития женских половых клеток (или дифференцировки половых клеток по женскому типу) называют оогенезом.
Структура оогенеза примерно одинакова у всех животных (рисунок 3.1). Гоноцит попадает в яичник и становится оогонием. Оогонии – незрелые половые клетки, способные к митозу. Оогенез совершается в 3 этапа (периода):
1) период размножения;
2) период роста;
3) период созревания.
Оогонии осуществляют первый период оогенеза – период размножения. Они делятся митотически, а затем переходят к стадии роста и становятся ооцитами I порядка. Они уже теряют способность к митотическому делению и вступают в профазу I мейоза. В период роста одновременно идут 2 процесса: в ядре происходит профаза мейоза, а в цитоплазме растущего ооцита I накапливаются органеллы, питательные вещества, источники энергии для зародыша. Вначале рост незначителен, и пропорционально увеличиваются объёмы ядра и цитоплазмы. В цитоплазме накапливаются РНК, белки, рибосомы, органеллы. Это малый рост (превителлогенез). Он совпадает с моментом вступления ооцита в профазу мейоза и продолжается до середины диплотены.
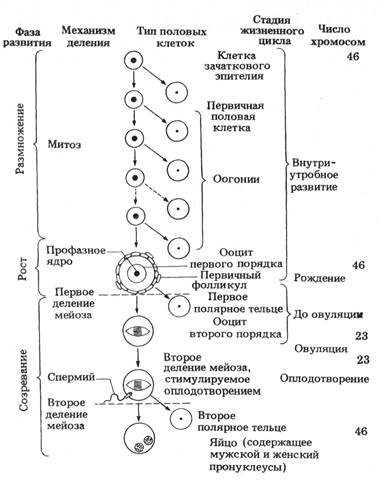
Рисунок 3.1 – Формирование яйцеклеток человека
Профаза I деления – это очень ответственный этап мейоза. И в оо-, и в сперматогенезе она включает стадии лептотены, зиготены, пахитены, диплотены и диакинеза. В лептотене реплицированные хромосомы деспирализуются и принимают вид длинных, спутанных нитей. В зиготене гомологичные по отцовской и материнской линиям хромосомы сближаются и конъюгируют (соединяются попарно), образуя биваленты. В пахитене хромосомы обмениваются генами и группами генов (происходит кроссинговер). Конъюгация – настолько важное событие в развитии организмов, что если он не произойдёт, например, при отсутствии гомологичной хромосомы, не образуются и гаметы. Именно поэтому оказываются бесплодными межвидовые гибриды с несовпадающими наборами хромосом. В диплотене гомологичные хромосомы, образующие бивалент, расходятся, но между ними сохраняется связь в области центромер.
Диакинез заключительная стадия профазы I деления мейоза. Хромосомы спирализуются и укорачиваются. Примерно с середины диплотены ооциты I вступают в период большого роста (вителлогенез). При этом резко увеличивается объём цитоплазмы (в сотни тысяч раз), и она постепенно заполняется желточными включениями. Желток – смесь белков, жиров и углеводов, основной из них – вителлогенин (синтезируется в печени).
Заключительный этап оогенеза – период созревания. Он осуществляется в виде двух последовательных делений созревания. В результате первого деления мейоза реплицированные гомологичные хромосомы расходятся в дочерние ядра, каждое из которых содержит половину исходного числа хромосом. В результате второго деления созревания в сестринских клетках оказывается гаплоидное число одиночных хроматид.
Особенностью делений созревания в оогенезе является асимметричное разделение цитоплазмы: при каждом делении почти вся она остаётся в одной из сестринских клеток. При первом делении образуются большой ооцит второго порядка (ооцит II) и маленькое первое редукционное тельце. В цитоплазме ооцита содержится практически всё, что накоплено в период роста. При втором делении созревания образуется зрелое яйцо с тремя редукционными тельцами, которые впоследствии обычно дегенерируют.
Существуют следующие типы оогенеза: диффузный и локализованный (рисунок 3.2). Диффузный оогенез: развитие женских гамет может осуществляться в любом участке тела (губки, кишечнополостные, ресничные черви). При диффузном типе ооциты не синтезируют и не накапливают желточные включения, а растут за счёт низкомолекулярных соединений из фаголизосом. Это фагоцитарный тип питания.
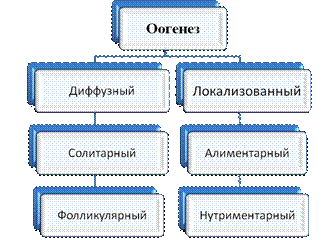
Рисунок 3.2 – Виды оогенеза
Локализованный оогенез: развитие женских гамет локализовано в личинках. При локализованном оогенезе ооцит может развиваться без участия вспомогательных питающих клеток (солитарный тип) или при участии специальных клеток (алиментарный тип).
Алиментарный тип оогенеза может быть фолликулярным или нутриментарным.
При фолликулярном оогенезе растущий ооцит окружен фолликулярными клетками и образует фолликул. Фолликулярные клетки не участвуют в синтезе белков желтка, все виды РНК синтезируются в самом ооците (у всех хордовых)
При нутриментарном оогенезе ооцит окружен трофоцитами (клетками-кормилками, абортивными половыми клетками).
Яйцеклетка – крупная и неподвижная, размеры её у млекопитающих 100-200 мкм. Она содержит запас трофических соединений, необходимых для раннего развития зародыша, а так же запас цитоплазматических структур для формирования бластомеров. Количество желтка варьирует у животных разных групп.
На ультрамикроскопическом уровне яйцеклетка имеет крупное округлое ядро с эухроматином и цитоплазму с митохондриями, рибосомами и полисомами. В цитоплазме есть окаймлённые везикулы (свидетельства активных транспортных процессов) и мультиламеллярные тельца (значение до конца не выяснено). Кроме того, в цитоплазме разбросаны немногочисленные скопления вителлиновых гранул (это коллекторы трофических включений).
У человека и у высших плацентарных млекопитающих женские гаметы не существуют изолированно. Кроме самых ранних стадий развития (овогонии), они находятся в тесном контакте с соматическими клетками яичника, которые формируют вокруг каждой яйцеклетки эпителиальную и соединительнотканную оболочки. Комплекс «половая клетка – соматические оболочки» именуется овариальным фолликулом. Его строение постепенно усложняется в процессе овогенеза.
Яйцевые оболочки – защитные образования различного строения, окружающие яйца у подавляющего большинства животных. Лишь у немногих животных, яйца которых способны к амебоидному движению, например у губок и некоторых кишечнополостных, поверхность яйца не защищена оболочкой. По происхождению различают три типа яйцевых оболочек.
Первичную, или желточную, оболочку вырабатывает само яйцо в период роста (ооцит). Нередко она представлена тонким прозрачным слоем, иногда (например, у аскариды, некоторых позвоночных) достигает значительной толщины и может быть многослойной. У большинства позвоночных желточная оболочка пронизана многочисленными радиальными каналами, за что получила название zona radiata; у млекопитающих её называют блестящей оболочкой — zona pellucida. Каналы возникают при формировании оболочки в тех местах, где располагаются отходящие от поверхности ооцита микроворсинки, направленные навстречу выростам окружающих его фолликулярных клеток. Некоторые исследователи полагают, что образование zona radiata осуществляется за счёт веществ, секретируемых не только ооцитом, но и фолликулярными клетками, поэтому её нельзя считать первичной в строгом смысле слова. Желточная оболочка образуется почти всегда; многие яйца имеют, кроме того, оболочку другого типа (обычно вторичную), а иногда — оболочки всех трёх типов.
Вторичная оболочка, или хорион, выделяется вспомогательными клетками яичника или формируется путём преобразования этих клеток в материал оболочки. Большой прочности достигает хорион у насекомых и других членистоногих, у которых он пропитывается хорионином — веществом, по составу близким кератину. Третичные оболочки секретируются клетками полового тракта самки во время продвижения овулировавшего яйца по яйцеводу.
К третичным оболочкам относятся студенистые оболочки яиц иглокожих, моллюсков, рыб и земноводных, а также более плотные белковые оболочки, одетые роговой скорлупой у головоногих моллюсков и акулообразных рыб, волокнистой и известковой у пресмыкающихся или подскорлуповыми оболочками и известковой скорлупой у птиц. У многих беспозвоночных прочная третичная оболочка окружает несколько яиц и называется яйцевым коконом. Кокон содержит белковую жидкость, которая является жидкой третичной оболочкой. Если плотные оболочки формируются до соединения яйца и сперматозоида, в них образуются особые каналы (микропиле), по которым оплодотворяющий сперматозоид может беспрепятственно достигнуть цитоплазмы яйца. Яйцевые оболочки защищают яйца от механических повреждений, от проникновения микроорганизмов и паразитов, от высыхания и т. п. Белковая оболочка куриного яйца обладает бактерицидными, фунгицидными свойствами. У многих животных клейкие оболочки или их придатки служат также для прикрепления яиц к субстрату. Материал жидких белковых оболочек используется развивающимися зародышами как питательный материал. Отделение оболочки от поверхности яйца при оплодотворении играет важную роль в его защите от проникновения сверхчисленных сперматозоидов.