 2015-04-30
2015-04-30 1137
1137Перекись водорода Участок молекулы жирной кислоты, подвергшейся СРО
Далее в этом месте происходит расщепление жирной кислоты.
В связи с возникновением перекисной группировки свободноради- кальное окисление жирных кислот, входящих в состав липоидов, часто обозначают термином перекисное окисление липидов (ПОЛ).
Образование свободных радикалов кислорода в тканях организма происходит постоянно. За счет этого процесса прежде всего осуществляется обновление липидного слоя биологических мембран.
Считается, что свободные радикалы кислорода выполняют также защитную функцию, окисляя различные чужеродные вещества, поступающие в организм извне, в том числе мембранные белки и липоиды патогенных микроорганизмов.
В физиологических условиях свободнорадикальное окисление протекает с низкой скоростью, так как ему противостоит защитная анти- оксидантная система организма, предупреждающая накопление свободных радикалов кислорода и ограничивающая тем самым скорость СРО. Главным компонентом антиоксидантной системы является витамин Е (токоферол).
Активные формы кислорода могут возникать в процессе тканевого дыхания. Для нормального течения тканевого дыхания необходимо одновременное присоединение к молекуле кислорода четырех электронов. В этом случае образуются две молекулы воды. Однако же иногда (например, при избытке кислорода) к молекуле кислорода присоединяются два или даже один электрон. В этих случаях вместо воды возникают соответственно перекись водорода (Н202) и супероксид-анион кислорода (02~), которые очень токсичны для клеток, так как являются сильными окислителями и повреждают биомембраны (подробно см. в главе 17 «Молекулярные механизмы утомления»).
Для защиты от этих опасных соединений в состав антиоксидантной системы входят специальные ферменты. Под действием фермента су- пероксиддисмутазы супероксиданион превращается в перекись водорода:
2 02~ + 2 Н+--------- Н202 + 02
Далее перекись водорода разрушается ферментом каталазой: 2 Н202 — 2 Н20 + 02
ГЛАВА 5 ОБМЕН УГЛЕВОДОВ
гткРНВАРИВАНИЕ И ВСАСЫВАНИЕ УГЛЕВОДОВ
С пищей в сутки поступает 400-500 г углеводов. Основные пищевые углеводы - крахмал, клетчатка, сахароза (пищевой сахар), лактоза (молочный сахар), гликоген.
Переваривание пищевых углеводов начинается в ротовой полости. Под действием фермента слюны амилазы крахмал и гликоген подвергаются неглубокому расщеплению с образованием низкомолекулярных полисахаридов - декстринов. Дальнейший распад декстринов, а также нерасщепленного крахмала и гликогена протекает в тонкой кишке с участием амилазы поджелудочного сока. В результате образуется диса- харид мальтоза, состоящая из двух остатков глюкозы. Завершается переваривание углеводов превращением образовавшейся мальтозы и других пищевых дисахаридов (сахароза, лактоза) в моносахариды (глюкоза, фруктоза, галактоза), главным из которых является глюкоза.
Клетчатка (целлюлоза), в молекуле которой остатки глюкозы соединены прочными связями, в ходе пищеварения не расщепляется и, пройдя через весь кишечник, выделяется из организма.
Образовавшиеся моносахариды всасываются по системе воротной вены и поступают вначале в печень. При этом в печень поступает практически только глюкоза, так как в ходе всасывания в клетках тонкой кишки в нее могут превращаться другие моносахариды (фруктоза, галактоза и др.).
В печени значительная часть глюкозы превращается в гликоген, который представляет собою запасную, резервную форму глюкозы или депо глюкозы (свободная глюкоза накапливаться в клетках не может, так как ее молекулы имеют малый размер и легко проходят через клеточные мембраны). Между приемами пищи в печени протекает противоположный процесс - гликоген распадается на глюкозу, которая из печени выходит в кровь.
СИНТЕЗ ГЛИКОГЕНА________________________________________________
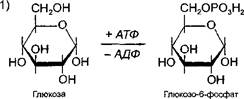
Глюкоза, используемая для синтеза гликогена, предварительно активируется. Вначале глюкоза взаимодействует с АТФ и превращается в ппокозо-6-фосфат, который затем легко переходит в глюкозо-1-фосфат (формулы этих соединений см. ниже в разделе «Метаболизм глюко- л
зы»), Далее глюкозо-1 -фосфат реагирует с УТФ (уридинтрифосфат - макроэргическое соединение, похожее по строению на АТФ и содержащее вместо аденина урацил). В ходе этой реакции отщепляются два остатка фосфорной кислоты в виде дифосфата и образуется очень активная форма глюкозы - уридиндифосфатглюкоза (УДФ-глюкоза),
Схематично активацию глюкозы можно представить следующим образом:
+ АТФ + УТФ
Гл -------- — Гл-6-ф-------- — Гл-1-ф ------ —— Гл-1-УДФ
-АДФ - фф
Синтез гликогена осуществляется путем присоединения образовавшейся УДФ-глюкозы к наружным цепям молекул имеющегося в клетках печени гликогена, который называется затравкой. При этом в молекулу гликогена включаются только остатки глюкозы. В результате многократного присоединения остатков глюкозы наружные цепи удлиняются и разветвляются, что ведет к значительному увеличению размера молекул гликогена.
Синтез гликогена может быть описан следующим уравнением:
(C6H10O5)n + m УДФ-С6Н10О5 ---------- (C6H10O5)n+m + m УДФ
Гликоген-«затравка» УДФ-тюкоза Удлиненный гликоген
Освобождающиеся в процессе синтеза гликогена молекулы УДФ вступают в реакцию с АТФ и снова превращаются в УТФ:
УДФ + АТФ ------- УТФ + АДФ
Таким образом, источником энергии для синтеза гликогена является АТФ, а УТФ выполняет роль переносчика энергии.
Благодаря синтезу в печени происходит накопление гликогена и его концентрация может достигать 5-6%. Превращение в печени глюкозы в гликоген предотвращает резкое увеличение ее содержания в крови во время приема пищи.
Синтез гликогена из глюкозы также происходит в мышцах, но его концентрация в них не превышает 2-3%. Образованию гликогена в мышцах способствует пищевая гипергликемия - повышение концентрации глюкозы крови во время приема пищи, обусловленное тем, что часть глюкозы проходит через печень в большой круг кровообращения. Кроме этого, незначительная часть глюкозы может всасываться из кишечника по лимфатической системе и, минуя печень, сразу попадать в большой круг кровообращения.
Синтез гликогена ускоряется гормоном инсулином.
РАСПАД ГЛИКОГЕНА
Между приемами пищи гликоген печени расщепляется и превращается в глюкозу, которая выходит в кровь. Этот распад идет с участием фосфорной кислоты и называется фосфоролизом. Под действием фосфорной кислоты от наружных цепей гликогена поочередно отщепляются остатки глюкозы в форме глюкозо-1-фосфата. Полностью гликоген не расщепляется. Оставшиеся небольшие молекулы гликогена служат в дальнейшем «затравкой» при его синтезе из глюкозы.
Фосфоролиз гликогена протекает по следующему уравнению: (C6H10O5)n + m Н3Р04 (CeHioOg)^ +тГл-1-ф
Исходный гликоген Гликоген-«затравка»
Последующее превращение глюкозо-1-фосфата в свободную глюкозу осуществляется в две стадии. На первой стадии глюкозо-1-фосфат переходит в глюкозо-6-фосфат; на второй стадии происходит гидролиз глюкозо-6-фосфата и образуется свободная глюкоза и фосфорная кислота:
+ н2о
Гл-1-ф ------- — Гл-6-ф ---------- — Глюкоза + Н3Р04
Распад гликогена в печени до глюкозы часто обозначается термином глюкогенез, он ускоряется гормонами глюкагоном и адреналином.
Благодаря протеканию в печени двух противоположных процессов: синтеза гликогена из глюкозы и его распада снова на глюкозу, ее концентрация в крови изменяется только в небольшом диапазоне, поэтому кровь постоянно снабжает все органы глюкозой.
В мышцах расщепление гликогена обычно наблюдается при выполнении физической работы. Однако свободная глюкоза здесь не образуется, так как в мышечных клетках нет фермента, вызывающего гидролиз глюкозо-6-фосфата. Глюкозо-1-фосфат и глюкозо-6-фосфат из-за наличия фосфатного остатка через стенку мышечных клеток проходить не могут, поэтому все дальнейшие превращения этих соединений протекают непосредственно в мышцах и направлены на обеспечение их энергией.
Распад гликогена в мышцах стимулирует гормон адреналин, который выделяется в кровь как раз во время мышечной работы.
МЕТАБОЛИЗМ ГЛЮКОЗЫ
Использование глюкозы в организме осуществляется двумя путями:
• Большая часть глюкозы (90-95%) подвергается распаду по гек- созодифосфатному пути (ГДФ-путь), который является для организма главным источником энергии.
• Незначительная часть глюкозы (5-10%) распадается по гексозо- монофосфатному пути (ГМФ-путь), имеющему анаболическое назначение и обеспечивающему различные синтезы рибозой и водородом в форме НАДФ-Н2.
Гексозодифосфатный путь (ГДФ-путь) ________________________________
ГДФ-путь может протекать аэробно и анаэробно. Аэробный ГДФ- путь функционирует постоянно, а анаэробный распад углеводов наблюдается только при повышенной потребности клеток в энергии, в основном в скелетных мышцах.
Аэробный распад углеводов
Аэробный распад углеводов по ГДФ-пути - сложный, многостадийный процесс, включающий десятки промежуточных реакций, приводящих в конечном счете к образованию углекислого газа и воды с выделением большого количества энергии. Этот процесс можно разделить на три этапа, последовательно идущих друг за другом.
Первый этап ГДФ-пути происходит в цитоплазме клеток. На этом этапе глюкоза превращается в пировиноградную кислоту (пируват). Этот этап часто называют гликолизом.
На первой стадии глюкоза путем взаимодействия с АТФ переходит в активную форму - глюкозо-6-фосфат:
|
Это единственная реакция, которой подвергается в организме глюкоза. Поэтому все превращения гчюкозы в организме начинаются с образования глюкозо-6-фосфата. Далее глюкозо-б-фосфат вступает б
различные пути метаболизма глюкозы (например, рассмотренный вы ше синтез гликогена).
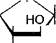
На следующих стадиях глюкозо-6-фосфат изомеризуется во фрукто- зо-6-фосфат, который, взаимодействуя с АТФ, далее превращается во (Ьг)уктозо-1,6-дифосфат. (Этим объясняется название данного пути распада углеводов - гексозодифосфатный путь, поскольку фруктоза содержит шесть атомов углерода и относится к гексозам.)
Перечисленные реакции можно описать следующими уравнениями:
2) СН20Р03Н2 1——О
СН2ОН ^ 2 + АТФ
но^Дн "" ^С
| 3) СН20Р03Н2 |
| ОН -АДФ НО ' Фруктозо-6-фосфат |
| он он Глюкозо-6-фосфат |
I I.. L1А
З1 '2
СН20Р03Н2
( ОН ОН
| СН90Р0,Н |
| о |
Фруктозо-1,6-дифосфат
Образовавшийся фруктозо-1,6-дифосфат расщепляется на две фос- фотриозы - фосфоглицериновый альдегид и фосфодиоксиацетон,
которые являются изомерами и легко переходят друг в друга:
12КУ1 V-f3l 12
,0. СН20Р03Н2
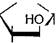
|
( ОН
он
| 4) СН,ОРО,Н, |
Фруктозо-1,6-дифосфат
С'" неон
I
СН20Р03Н2
Фосфоглицериновый альдегид
СН2ОН
I
с=о
I
СН20Р03Н2
Фосфодиоксиацетон
В последующих реакциях данного этапа участвует только фосфоглицериновый альдегид, и по мере его использования в него превращается фосфодиоксиацетон:
— неон
I
СН20Р03Н2
| 5) |
| СН2ОН I с=о I СН20Р03Н2 Фосфодиоксиацетон |
Фосфоглицериновый альдегид
Поэтому можно считать, что из глюкозы образуется две молекулы Фосфоглицеринового альдегида.
Следующая стадия - окисление фосфоглицеринового альдегида, протекающее непосредственно в цитоплазме. В ходе этой реакции от окисляемого вещества отнимаются два атома водорода и временно присоединяются к коферменту НАД. За счет выделяющейся при окислении энергии в продукт реакции включается еще один фосфатный остаток, который присоединяется макроэргической связью:
6) /Н ^О
I ^О I ^0~Р03Н2
неон + НАД + Н3Р04 —- неон + НАДН2
СН20Р03Н2 СН2ОРОэН2
Фосфоглицериновый альдегид 1,3-дифосфоглицерат
При невысокой скорости распада углеводов (в покое или при работе умеренной мощности) весь образовавшийся НАД-Н2 передает атомы водорода в дыхательную цепь митохондрий, где эти атомы связываются с молекулярным кислородом и превращаются в воду. За счет выделяющейся при этом энергии осуществляется синтез АТФ. Как уже отмечалось (см. главу 4 «Биологическое окисление»), перенос двух атомов водорода на кислород сопровождается синтезом трех молекул АТФ.
Таким образом, в данных условиях первый этап ГДФ-пути протекает аэробно. Поскольку из глюкозы образуется две молекулы фосфоглицеринового альдегида и соответственно две молекулы восстановленного НАД, то в расчете на одну молекулу глюкозы в процессе тканевого дыхания осуществляется синтез шести молекул АТФ.
На следующей стадии фосфатный остаток, благодаря наличию макроэргической связи, легко передается на молекулу АДФ с образованием АТФ:
7)
С С
|^0~Р03Н2 |^ОН
НСОН + АДФ ---------- — НСОН + АТФ
I I
СН20Р03Н2 СН20Р03Н2
1,3-дифосфоглицерат 3-фосфоглицерат
Такой способ синтеза АТФ, осуществляющийся без участия тканевого дыхания и, следовательно, без потребления кислорода, обеспеченный запасом энергии субстрата, называется анаэробным, или субстратным, фосфорилированием. Это самый быстрый путь получения АТФ.
На последующих стадиях образовавшийся 3-фосфоглицерат изоме- изуется в 2-фосфоглицерат, от которого затем отщепляется молекула воды что приводит к перераспределению энергии в молекуле и возникновению макроэргической связи:
я\ 9)
СООН СООН СООН
I I I
НСОН ------------ — НС0Р03Н, ----------- — С-0~Р03Н2
| I -Н20 II
СН20Р03Н2 СН2ОН сн2
3-фосфоглицерат 2-фосфоглицерат Фосфоенолпируват
Завершается первый этап ГДФ-пути реакцией анаэробного фосфо- рилирования, в ходе которой синтезируется еще одна молекула АТФ:
10) СООН СООН I I С-0~Р03Н2 +АДФ — С = 0 + АТФ
СН2 СН2
Фосфоенолпируват Пируват
(Пировиноградная кислота)
Учитывая, что из одной молекулы глюкозы образуется две молекулы фосфоглицеринового альдегида, всего синтезируется десять молекулы АТФ (шесть - аэробно и четыре - анаэробно). При этом следует учесть, что на первых стадиях расходуется две молекулы АТФ на активацию глюкозы и фруктозо-6-фосфата. В итоге превращение глюкозы в пируват сопровождается синтезом восьми молекул АТФ.
Суммируя уравнения всех стадий, можно получить итоговое уравнение первого этапа:
С6Н12Ое + 02 + 8 АДФ + 8 Н3Р04 -------- — 2 С3Н403 + 2 Н20 + 8 АТФ
Глюкоза Пируват
Первый этап распада углеводов практически обратим. Из пирувата, а также из лактата (см. ниже) может синтезироваться глюкоза, а из нее затем гликоген.
Второй и третий этапы ГДФ-пути протекают в митохондриях с участием дыхательной цепи и поэтому обязательно требуют 02. Эти этапы, в отличие от первого, необратимы.
В ходе второго этапа от пировиноградной кислоты отщепляется углекислый газ и два атома водорода. Отщепленные атомы водорода по дыхательной цепи передаются на кислород с выделением воды и одновременным синтезом АТФ, а образовавшаяся из пирувата уксусная кислота присоединяется макроэрги ческой связью к коферменту А - переносчику остатков кислот.
Кофермент А содержит в своей молекуле остаток витамина В,- пантотеновой кислоты — и имеет свободную SH-группу. В сокращенном виде он обозначается - HSKoA.
Образовавшийся комплекс уксусной кислоты и кофермента А называется ацетилкофермент А. Уксусная кислота, связанная с кофермен- том А, обладает высокой химической активностью, поэтому ацетилкофермент А часто называют активной уксусной кислотой.
В одной из реакций этого этапа еще участвует в качестве кофермента производное витамина В, - тиаминдифосфат.
Итоговое уравнение второго этапа ГДФ-пути: 03Н403 + 1/2 02 + HSKoA + 3 АДФ + 3 Н3Р04 —
Пируват Кофермент А
О
СН3 - С ~ SKoA + С02 + Н20 + 3 АТФ
Ацетил-КоА
На третьем этапе остаток уксусной кислоты, входящий в состав ацетилкофермента А, подвергается дальнейшему окислению и превращается в С02 и Н20. Этот этап носит циклический характер и называется циклом трикарбоновых кислот (ЦТК), или циклом Кребса. За счет выделяющейся энергии на этом этапе также осуществляется синтез АТФ.
Цикл трикарбоновых кислот (ЦТК) - это завершающий этап катаболизма не только углеводов, но и всех остальных классов органических соединений. Это обусловлено тем, что при распаде углеводов, жиров и аминокислот образуется общий промежуточный продукт - уксусная кислота, связанная со своим переносчиком - коферментом А - в форме ацетилкофермента А.
Вышесказанное можно иллюстрировать следующей схемой:
Белки Углеводы Жиры (аминокислоты)^\^
Ацетил-КоА \ЦТК

|
С02 н2о
Цикл Кребса протекает в митохондриях с обязательным потреблением кислорода и требует функционирования тканевого дыхания.
На первой стадии цикла остаток уксусной кислоты переносится с молекулы ацетилкофермента А на молекулу щавелево-уксусной кислоты (ЩУК) с образованием лимонной кислоты:
СН3 + I
С = О ^SKoA
| СООН I СН, I с=о I СООН щук |
| + Н20 |
Ацетил-КоА
СООН I
сн2
I
НО-С-СООН + I
сн2
I
СООН
Лимонная кислота
HS - КоА
Кофермент А
Лимонная кислота содержит три карбоксильные группы, т. е. является трикарбоновой кислотой, что обусловило название этого цикла.
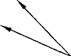
Далее, от лимонной кислоты поочередно отщепляются две молекулы С02 и четыре пары атомов водорода и вновь образуется ЩУК (в связи с этим рассматриваемый процесс называется циклом). Отщепленный водород по дыхательной цепи передается на молекулярный кислород с образованием воды. Перенос каждой пары атомов водорода на кислород сопровождается синтезом трех молекул АТФ. Всего при окислении одной молекулы ацетилкофермента А синтезируется 12 молекул АТФ.
Итоговое уравнение цикла Кребса (третьего этапа ГДФ-пути):
О
СН3 - С ~ SKoA + 2 02 + 12 АДФ + 12 Н3Р04 --------------- —
Ацетилкофермент А
HSKoA + 2 СО, + Н,0 + 12 АТФ
Схематично цикл Кребса можно представить следующим образом:
| HSKoA |
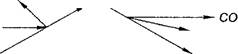
| Ацетил-КоА (С2) |
| Щавелево-уксусная кислота (С4) |
| 2 Н (+ 1/2 Ог, -ЗАТФ) |
| Яблочная кислота (С4) |
Лимонная кислота (С6)
2 Н(+ 1/2 Ог, -ЗАТФ) а-кетоглутаровая кислота (С5)
Г^ с°2
2 Н (+ 1/2 02, -ЗАТФ)
Янтарная кислота (С4)
2 Н (+ 1/2 02, - 3 АТФ)
Биологическая роль цикла трикарбоновых кислот заключается в том, что он является главным источником АТФ для организма. Цикл Кребса дает АТФ больше, чем все вместе взятые предшествующие ему процессы образования ацетилкофермента А из белков, углеводов и жиров.
Суммируя уравнения всех трех этапов, можно получить итоговое уравнение аэробного ГДФ-пути окисления глюкозы в целом (коэффициенты в уравнениях второго и третьего этапов необходимо удвоить, так как из одной молекулы глюкозы образуется две молекулы пирувата и соответственно две молекулы ацетил-КоА):
Итоговое уравнение аэробного ГДФ-пути: С6Н120в + 6 02 + 38 АДФ + 38 Н3Р04 6 С02 + 6 Н20 + 38 АТФ
Анаэробный распад углеводов
Анаэробный распад углеводов обычно протекает в мышцах при выполнении интенсивных нагрузок. По своему содержанию анаэробный распад соответствует первому этапу ГДФ-пути. Однако из-за высокой скорости этого процесса образующиеся в больших количествах НАД-Н2 и пируват не полностью окисляются в митохондриях. В этом случае большая часть НАД-Н2 передает атомы водорода пировиноградной кислоте непосредственно в цитоплазме:
СООН СООН
I I
С = 0 + НАД-Н2 —— СН-ОН + НАД сн3 сн3
Пируват Лакгат
В этом случае все реакции протекают анаэробно (без участия митохондрий и потребления кислорода) и приводят к образованию и накоплению лактата (молочной кислоты). Такой анаэробный распад углеводов получил название анаэробный гликолиз, или просто гликолиз.
Гликолиз полностью протекает в цитоплазме клеток, и синтез АТФ происходит только анаэробно (см. 7-ю и 10-ю стадии первого этапа аэробного ГДФ-пути).
В мышцах гликолизу в основном подвергается гликоген (его в мышцах много!), и его анаэробному распаду соответствует следующее итоговое уравнение:
(С6Н10О5)п + 3 АДФ + 3 Н3Р04---------- (С6Н10О5)п„, + 2 С3Н603 + 3 АТФ
Гликоген Укороченный Лактат
гликоген
Анаэробный распад гликогена является дополнительным способом получения АТФ при выполнении интенсивной мышечной работы, протекающим без участия митохондрий и потребления кислорода.
Анаэробный распад глюкозы наблюдается главным образом в красных клетках крови (эритроцитах), в которых отсутствуют митохондрии. Для этих клеток гликолиз является основным источником энергии.
Итоговое уравнение гликолитического распада глюкозы следующее:
С6Н1206 + 2 АДФ + 2 Н3Р04 --------------------------------- 2 С3Н603 + 2 АТФ
Глюкоза Лактат
Аэробный и анаэробный распад глюкозы и гликогена протекают практически одинаково, но при любом распаде гликогена образуется на одну молекулу АТФ больше, так как в этом случае образование глюко- зо-6-фосфата происходит без использования АТФ (см. с. 39).
В целом ГДФ-путь распада углеводов может быть представлен следующей упрощенной схемой:
Глюкоза Гликоген
Гл-6-ф
I
Пируват
Ацетил-КоА
I
УЧ
со2 н2о
(+ 38 АТФ при распаде глюкозы; + 39 АТФ при распаде гликогена в расчете на одну молекулу глюкозы)
ч
Лактат
(+ 2 АТФ при распаде глюкозы; +■ 3 АТФ при распаде гликогена в расчете на одну молекулу глюкозы)
Соотношение между аэробным и анаэробным распадом углеводов определяется потребностью клеток в энергии (АТФ). При низкой и средней потребности клеток в АТФ преобладает аэробное окисление, а при высокой потребности в энергии большая часть углеводов превращается в молочную кислоту, т. е. используется в гликолизе.
£&$£оз омонофосфатный путь распада углеводов (ГМФ-пут ь)
Распаду по ГМФ-пути в организме подвергается незначительная Часть глюкозы (5-10%). Данный путь распада в основном встречается в "ечени, надпочечниках, эритроцитах, жировой ткани и протекает в ци- т°плазме клеток.
ГМФ-путь распада глюкозы имеет анаболическое назначение и обеспечивает различные синтезы рибозой и водородом в форме НАДФН2.
ГМФ-путь можно разделить на два этапа, причем первый этап протекает обязательно, а второй не всегда.
Первый этап начинается с перехода глюкозы в активную форму - глюкозо-6-фосфат, от которого затем отщепляется молекула углекислого газа и две пары атомов водорода, присоединяющиеся к коферменту НАДФ (никотинамидадениндинуклеотидфосфат). По строению НАДФ похож на НАД, но, в отличие от последнего, имеет дополнительную фосфатную группу.
Конечным продуктом первого этапа является рибозо-5-фосфат (моносахарид, содержащий пять атомов углерода и связанный с остатком фосфорной кислоты, т. е. пентозофосфат).
Итоговое уравнение первого этапа:
2 НАДФ 2 НАДФ Н2
+АТФ V У
Гл------ — Гл-6-ф ^—Риб-5-ф
-АДФ -С02
Образовавшийся НАДФ-Н2 поставляет атомы водорода в различные синтетические процессы, в том числе для синтеза жирных кислот и холестерина. Рибозо-5-фосфат используется для синтеза нуклеотидов, из которых далее образуются нуклеиновые кислоты и коферменты (например, кофермент А, НАД, НАДФ, ФМН, ФАД).
Второй этап протекает тогда, когда рибозо-5-фосфат полностью не расходуется для синтезов. Неиспользованные молекулы этого вещества вступают во взаимодействия друг с другом, в ходе которых они обмениваются группами атомов и в качестве промежуточных продуктов появляются моносахариды с различным числом атомов углерода (триозы, тетрозы, пентозы, гептозы). В конечном счете из шести молекул рибозо-5-фосфата образуется пять молекул глю- козо-6-фосфата:
6 Риб-5-ф ----------- — 5 Гл-6-ф
Таким образом, благодаря второму этапу данный распад глюкозы приобретает циклический характер и его часто называют пентозным циклом.
Объединив оба этапа, можно получить общую схему пентозного
цикла:
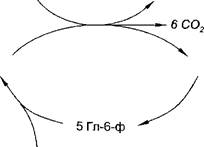
|
| 12 НАДФ |
| 6 Гл-6-ф |
| 12 НАДФ Н, |
| 6 Риб-5-ф |
| Гл-6-ф |
Полезная информация
Цикл трикарбоновых кислот (ЦТК) - центральный метаболический процесс организма - был обнаружен и изучен крупнейшим биохимиком XX века Г. Кребсом.
За это открытие Г. Кребсу была присуждена Нобелевская премия.
ГЛАВА 6 ОБМЕН ЖИРОВ

|
В суточном рационе обычно содержится 80-100 г жиров. Переваривание жиров происходит в тонкой кишке. Жир предварительно с помощью желчных кислот превращается в эмульсию, что значительно повышает эффективность действия фермента поджелудочного сока - липазы. Под действием этого фермента жир расщепляется на глицерин и жирные кислоты: о
сн2 — о — С — R,
i о
1 II
сн — о — с — r2
i о
Ju 11
сн2 — о — с — r3
| + 3 Н,0 |
Жир
сн2он r, — соон
сн — он + r2—соон
r3— соон
Жирные кислоты
Глицерин
Поскольку в пище присутствуют разнообразные жиры, то в результате их переваривания образуется большое количество разновидностей жирных кислот. Продукты расщепления жира всасываются слизистой тонкой кишки. Глицерин растворим в воде, поэтому его всасывание происходит легко. Жирные кислоты, нерастворимые в воде, всасываются в виде комплексов с желчными кислотами. В клетках тонкой кишки глицерин вновь соединяется с жирными кислотами, но только с теми, которые входят в состав жиров организма человека. В результате синтезируется человеческий жир.
Вновь образовавшийся (ресинтезированный) жир по лимфатическим сосудам, минуя печень, поступает в большой круг кровообращения и откладывается в запас в жировых депо. Главные жировые депо организма - подкожная жировая клетчатка, большой и малый сальники, околопочечная капсула.
КАТАБОЛИЗМ ЖИРОВ
Использование жира в качестве источника энергии начинается с его выхода из жировых депо в кровяное русло. Этот процесс называется мобилизацией жира. Мобилизация жира ускоряется под влиянием симпатической нервной системы и гормона адреналина.
Основные превращения жира происходят в печени, где имеются активные ферменты жирового обмена.
В печени жир прежде всего подвергается гидролизу и превращается, так же как и в кишечнике, в глицерин и жирные кислоты.
Образовавшийся глицерин легко переходит в фосфоглицерино- вый альдегид, который является также промежуточным продуктом распада углеводов и поэтому вовлекается в углеводный обмен.
Жирные кислоты, являясь веществами химически неактивными, вначале активируются с использованием энергии АТФ и связываются со своим переносчиком - коферментом А (комплекс жирная кислота - кофермент А называется ацилкофермент А):
О
R—СООН + HSKoA
| + АТФ -АМФ -ФФ |
Жирная кислота
II








