 2015-05-13
2015-05-13 1424
1424| Типы растительности | Среднее годовое потребление радиации на фотосинтез, % от | Среднее годовое связывание солнечной энергии в нетто-продукции, % от | ||
| суммарной радиации | ФАР | суммарной радиации | ФАР | |
| Дождевые тропические леса | 1,5 | 4,5 | 0,6 | 1,5 |
| Летнезеленые лиственные леса | 0,6 | 1,6 | 0,4 | 1,0 |
| Бореальные хвойные леса | 0,5 | 1,1 | 0,3 | 0,8 |
| Тропические травяные сообщества | 0,2 | 0,6 | 0,2 | 0,5 |
| Злаковники умеренной зоны | 0,2 | 0,6 | 0,2 | 0,5 |
| Тундры | 0,2 | 0,4 | 0,1 | 0,2 |
| Полупустыни | 0,02 | 0,05 | 0,02 | 0,04 |
| Сельскохозяйственные земли | 0,3 | 0,7 | 0,2 | 0,6 |
В процессе дыхания продуцентов, консументов и редуцентов и разложения органических остатков использованная при фотосинтезе энергия снова превращается в тепло, так что почти вся энергия, связанная первичными продуцентами, рассеивается и в отличие от вещества уже не возвращается в биологический цикл. При переходе от одного трофического уровня к другому на каждом последующем уровне для создания биомассы используется лишь небольшая часть энергии, заключенной в предыдущем уровне, и происходят ее большие потери.
По исследованиям в широколиственных лесах Западной Европы, из общего количества ассимилируемой энергии 54,7% расходуется на дыхание растительного покрова. Из 45,3%, накопленных в чистой первичной продукции, 18,9% остается в приросте, 1,1% выедается животными, 13,6% уходит в подстилку, 11,7% — в отмершие корни.
При разложении (дыхании) подстилки теряется еще 9,4%; 15,8% переходит в гумус (11,7% за счет корней и 4,1% за счет подстилки), но из них 14,0% уходит на «дыхание» (минерализацию гумуса) и лишь 1,9% накапливается. Всего, таким образом, на дыхание уходит 79,2% ассимилированной энергии и
20,8% удерживается в живом и мертвом органическом веществе.
Несколько иную структуру имеет модель трансформации энергии в
южнотаежных ельниках, построенная Ю. К. Шуйцевым (рис. 40) 1.
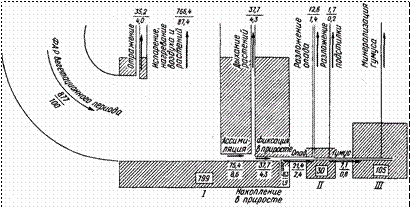 Рис. 40. Потоки энергии в южиотаежиом ельнике кисличнике (по материалам
Рис. 40. Потоки энергии в южиотаежиом ельнике кисличнике (по материалам
Ю. К. Шуйцева).
Основные блоки: I — фитомасса, II — мертвое органическое вещество (наземный и подземный опад), III
— почвенный гумус. Площади заштрихованных прямоугольников пропорциональны запасам энергии в блоках (Мдж/м2, обозначены цифрами в рамках). Эпюры со стрелками указывают направление основных потоков энергии; цифры в числителе — величина потока (Мдж/м2), в знаменателе — процент от количества ФАР вегетационного периода
В живой биомассе суши аккумулировано примерно 4*1016МДж энергии,
что соответствует,5% годовой суммарной солнечной радиации или около 14% радиационного баланса. В отдельных сообществах эти соотношения более высоки. Так, в таежных темнохвойных лесах запас связанной энергии составляет в среднем около 500 МДж/м2(что эквивалентно 40% годового радиационного баланса), в листопадных широколиственных — 650 МДж/м2(40%),
1См.: Шуйцев Ю. К. Некоторые особенности энергетики южнотаежных ельников Новгородской области//
Вопросы географии. 1981. № 117. С. 208 — 215.
в экваториальных лесах — 850 МДж/м2(24%), а в североамериканских лесах
из секвойи и дугласии — более 1700 МДж/м2(70%).
Некоторая часть аккумулированной солнечной энергии содержится в мертвом органическом веществе (подстилке, почвенном гумусе, торфе). В гумусе мощных тучных черноземов она превышает 1000 МДж/м2, в торфе — тысячи МДж/м2.
Ежегодно лишь ничтожная часть биологически связываемой энергии Солнца «консервируется» в мертвой органической массе и превращается в потенциальную химическую энергию, однако кумулятивный эффект этого процесса, продолжавшегося на протяжении геологических эпох, выразился в накоплении огромного запаса энергии в каустобиолитах, соразмерного с годовым поступлением суммарной солнечной радиации к суше.
Энергетическое значение биоты в функционировании геосистем определяется не только преобразованием солнечного тепла непосредственно при фотосинтезе (и разложении биомассы). Напомним, что уже на входе в
геосистему поток солнечной энергии существенно трансформируется
растительностью: низкое альбедо растительного покрова влияет на увеличение радиационного баланса; с другой стороны, под полог леса проникает лишь часть радиации (в темно-хвойном лесу — 1/10 и менее), причем изменяется ее спектральный состав и уменьшается доля ФАР;над поверхностью леса турбулентный поток тепла возрастает по сравнению с безлесной поверхностью и т. д. Особенно же существенное место в энергетическом балансе ландшафта занимают затраты солнечного тепла на перекачивание влаги из почвы в атмосферу через растения. В гумидных ландшафтах транспирация составляет главную статью расхода солнечной энергии. Как уже отмечалось, при развитом растительном покрове испарение осуществляется в основном в виде транспирации, за исключением неизбежных потерь влаги на физическое испарение (преимущественно до начала и после окончания вегетации)1.
Необходимость больших затрат энергии на транспирацию (и тем самым обеспечения минерального питания растений, а также предотвращения их перегрева), очевидно, служит причиной низкого КПД фотосинтеза и лимитирует возможности повышения биологической продуктивности. В ландшафтах, достаточно обеспеченных влагой, на транспирацию расходуется не менее 60 — 80% энергии радиационного баланса, а в экваториальных лесах этот показатель местами приближается к 100%. Если допустить гипотетическую возможность полного перехвата стока и физического испарения и утилизации всех атмосферных осадков, то в гумидных ландшафтах попросту не хватит солнечной энергии, чтобы растения смогли
«перекачать» все это количество влаги в атмосферу. В аридных же
ландшафтах
1Испарение происходит и с поверхности снежного покрова, составляя, впрочем, небольшую долю
суммарной эвапотранспирации — порядка 5 — 10 мм, или около 10 — 20% твердых осадков.
естественные пределы фотосинтеза и продуцирование биомассы определяются
недостатком осадков.
Топологические, или ландшафтно-морфологические (внутриландшафтные), закономерности в энергетике геосистем изучены недостаточно. Ю. К. Шуйцев, изучая энергетику южнотаежных ельников, пришел к заключению, что автономные (плакорные) сообщества используют солнечную энергию наиболее эффективно, а супераквальные (с застоем влаги)
— наименее. Трансэлювиальные и трансаккумулятивные местоположения занимают в этом отношении промежуточное место. Соответственно изменяются запасы энергии в живой фитомассе (МДж/м2): в плакорном ельнике-черничнике — 799, в чернично-зеленомошном и хвощево- папоротниковом — по 705, в сфагновом — 275. Запасы энергии в мертвом органическом веществе, напротив, возрастают по мере ухудшения дренажа и аэрации: в сухостое и подстилке соответственно 30, 77, 41, 204, а в гумусовом веществе почвы и торфе — 105, 99, 242 и 2396 МДж/м2.
Особый аспект энергетики ландшафта связан с потоками механической энергии. Источники механического перемещения вещества в ландшафте имеют двоякую природу: оно осуществляется за счет энергии тектонических процессов, «законсервированной» в земной коре, и энергии солнечных лучей. В «надводной» части материков (лежащей выше уровня Мирового океана) накоплено около 3*1018 МДж потенциальной механической энергии (примерно в 3 раза больше суммарной радиации, поступающей ежегодно на сушу), которая эквивалентна энергии тектонических процессов, затраченной на преодоление силы тяжести. Ежегодно при денудации реализуется, т. е. превращается в кинетическую энергию, около одной десятимиллионной доли
этого запаса, что соответствует десятитысячным долям процента от величины суммарной радиации. При кажущейся незначительности этой цифры с ней связаны мощные потоки твердого материала, о которых уже речь шла выше.
В количественном отношении на 2 — 3 порядка выше потоки механической энергии, происходящие за счет трансформации солнечного тепла и обусловливающие перемещения воздушных и водных масс, а также ледников, пыли, органического опада. Нарушение теплового и связанного с ним гравитационного равновесия в атмосфере из-за неравномерного поступления тепла от земной поверхности приводит к механическому перемещению воздушных масс, а вместе с ними — водяного пара, пылевых и органических частиц. В механическую энергию ветра ежегодно переходит n* 1014МДж солнечной энергии (около 0,1% суммарной радиации, полученной всей сушей). Эта энергия рассеивается в виде тепла (в том числе и при выпадении атмосферных осадков). Механическая энергия всех текучих вод, которая есть также не что иное, как трансформированная лучистая энергия Солнца, оценивается в n* 1013МДж*год (около 0,01% суммарной радиации).
В каждом конкретном случае запасы потенциальной механиче-
ской энергии геосистемы и величина работы, производимой при механическом перемещении вещества, могут быть определены путем сравнения потенциалов силы тяжести («энергия положения») масс, расположенных на разных гипсометрических уровнях. Например, если при разнице высот в 20 м в пределах площади 1 га годовой слой денудации составляет 0,1 мм при объемной массе 2,5 г/см3, т. е. перемещается 2,5*103кг материала, то работа по его перемещению (денудации) составит: Ад= 2,5*103кг * 9,8 м/с2* 20 м = 4,9 МДж/га = 4,9 • 10-4МДж/м2.
При годовом слое стока 300 мм с той же площади работа, произведенная текучими водами, равна: Ас= 3*106кг * 9,8 м/с2* 20 м = 59 МДж/га = 5,9 * 10-3МДж/м2.
Преобразование энергии может служить одним из показателей интенсивности функционирования ландшафта. Вопрос о возможности интегральной количественной оценки функционирования ландшафтов с помощью какой-либо единой меры имеет дискуссионный характер. Однако сравнивая ландшафты по отдельным частным показателям функционирования (трансформация солнечной энергии, внутренний влагооборот, биологический круговорот веществ и др.), мы находим между этими показателями определенное соответствие. Очевидно, интенсивность функционирования ландшафта тем выше, чем интенсивнее в нем внутренний оборот вещества и энергии и связанная с ним созидающая функция, которая выражается прежде всего в биологической продуктивности. В свою очередь, все перечисленные процессы определяются соотношением теплообеспеченности и увлажнения. Однако до сих пор не найдено удовлетворительного способа выражения этого соотношения в виде того или иного коэффициента. Наиболее удачным представляется предложенный Н. Н. Ивановым «показатель биологической эффективности климата» (ТК) в виде произведения от умножения суммы температур за период со средними суточными температурами выше 10 °С (Т) на годовой коэффициент увлажнения (К). При этом за предельную величину К принята 1, так как дальнейшее увеличение увлажнения не оказывает положительного эффекта на биологическую продуктивность и на функционирование ландшафта в целом.
При всей условности этого коэффициента (основной недостаток его состоит в том, что он «не работает» в приполярных областях, где не бывает средних температур выше 10 °С) он дает достаточно объективное представление об относительной степени тепло- и влагообеспеченности различных ландшафтов и при этом хорошо коррелирует с другими показателями функционирования, в качестве которых в табл. 15 приведены также относительные значения интенсивности внутреннего влагооборота, или, что то же, затрат тепла на эвапотранспирацию Е, величины чистой первичной продукции фитомассы Б и годового потребления зольных элементов и азота растительностью MN. По всем перечисленным показателям первое место занимают экваториальные ландшафты, для которых значения ТК, Е,
Т а б л и ц а 15. Показатели относительной интенсивности








