 2015-05-13
2015-05-13 1637
1637Открывший мутации в начале ХХ века голландский ботаник и генетик Х. Де Фриз (1848-1935) ошибочно полагал, что в результате крупных мутаций виды возникают сразу (скачкообразно), без естественного отбора. Однако дальнейшие исследования показали, что большинство крупных мутаций вредны. Поэтому многие учёные начали склоняться к тому, что мутации не могут служить материалом для эволюции.
Лишь в 20-х гг. ХХ века отечественные учёные С.С. Четвериков (1880-1956) и Н.И. Шмальгаузен (1884-1963) обосновали роль мутаций в эволюции. Было установлено, что любая природная популяция насыщена разнообразными мутациями. Последние чаще всего рецессивны и, находясь в гетерозиготном состоянии, не проявляются фенотипически. Такие мутации чаще всего и служат генетической основой эволюции. При скрещивании гетерозиготных особей эти мутации могут переходить у потомков в гомозиготное состояние и проявляться в фенотипе. Из поколения в поколение сохраняются особи с полезными мутациями, которые поддерживаются естественным отбором. Вредные мутации накапливаются в популяции в скрытом виде, создавая резерв изменчивости, что ведёт к изменению генофонда популяции. Резерв наследственной изменчивости увеличивается в результате перекомбинирования аллельных генов (комбинативной изменчивости).
Мутацию определяют как возникновение наследуемого изменения; в широком смысле это понятие приложимо как к отдельным генам, так и к целым хромосомам. Причинами индуцированных мутаций могут являться естественные и искусственные мутагенные факторы (рис. 143). В результате мутации возникают мутантный ген или мутантная хромосома, дающие начало мутантному признаку. Генные мутации могут возникнуть в любой момент, но их появление более вероятно в делящейся, а не в «покоящейся» клетке; при гаметогенезе появление мутаций наиболее вероятно во время мейоза. В генетическом отношении важны именно те мутации, которые возникают при гаметогенезе. Мутантный ген воспроизводится с высокой точностью до тех пор, пока он не изменится в результате новой мутации. В результате мутаций формируется элементарный эволюционный материал. Он используется далее в ходе эволюции как исходная основа для действия других элементарных эволюционных факторов.
И.И. Шмальгаузен пришёл к выводу, что практически любые мутации, нарушая нормальную сбалансированность процессов и функций в организме, вредны для организма, неблагоприятно влияют на фенотип. Поэтому поддерживаются отбором лишь их гетерозиготные состояния, благодаря чему рецессивные аллели могут накапливаться в популяции и «подгоняться» в ряду её поколений к старому генному комплексу. Очень редко возникают мутации, улучшающие фенотип, повышая его приспособленность к условиям среды. Однако именно они дают основной материал для естественного отбора.
|
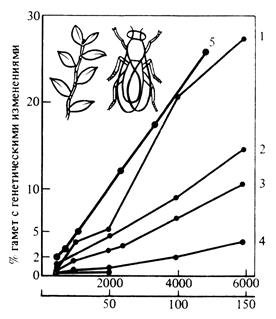
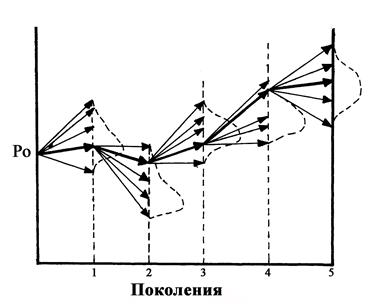
Мутационный процесс протекает в целом очень медленно и, соответственно, обусловливает медленную перестройку генетической структуры популяции. Наряду с мутациями на генетическую структуру популяции оказывают влияние другие процессы, в том числе дрейф генов (рис. 144). Дрейф генов - это изменение частоты генов в популяции в ряду поколений под действием случайных (стохастических) факторов. Он приводит, как правило, к снижению наследственной изменчивости популяций. Термин «дрейф генов» введён в 40-х годах ХХ века американским генетиком С. Райтом (1889-1982). Независимо от него Д.Д. Ромашов (1899-1963) и Н.П. Дубинин (1907-1998) описали это явление под названием «генетико-автоматический процесс». Дрейф генов проявляется наиболее отчётливо при резком сокращении численности популяции в результате стихийных бедствий (лесной пожар, наводнение), массового распространения вредителей и т.д.
Характерная особенность динамики генотипической структуры популяции под влиянием дрейфа генов заключается в:
а) усилении процесса гомозиготизации организмов, который нарастает с уменьшением численности популяции. Это нарастание обусловлено тем, что в популяциях ограниченного размера увеличивается частота близкородственных скрещиваний. Маловероятно, но тем не менее возможно, что некоторые из выщепившихся гомозиготных форм окажутся в новых условиях среды приспособительно ценными и смогут выжить, а не устраниться (элиминироваться) отбором;
|
б) потере популяцией некоторых аллелей и закреплении других, что особенно проявляется при значительном размахе колебаний численности в последовательных поколениях.
Вторым явлением, ускоряющим перестройку генофондов популяций, является миграция (поток) генов, возникающая при перемещении особей из одной популяции в другую и скрещивании с представителями последней. Если у пришельцев исходные частоты аллелей другие, чем у старожилов, то частоты аллелей во второй популяции могут измениться. Перемещение особей из популяции-донора в популяцию-реципиент приводит к изменению частот аллелей в обеих популяциях.

