 2015-05-10
2015-05-10 1066
1066В основе воспроизведения (репликации) структуры ДНК лежит так называемый принцип комплементарности: в двойной спирали две полимерные цепи ДНК связаны бок о бок водородными связями за счет образования пар G-C, C-G, А-Т и Т-А (см. [6], рис. 2 и 3). Если две цепи двойной спирали расходятся, то на каждой из них может строиться (полимеризоваться) новая комплементарная цепь, так что напротив G исходной цепи установится С новой цепи, напротив С старой цепи - G новой цепи, напротив А-Т, а напротив Т-А; в результате получатся две дочерние двойные спирали, полностью идентичные исходной - материнской (см. [6], рис. 4).
РНК химически подобна ДНК. В обоих случаях это линейные, неразветвленные полимеры нуклеотидов с пентозо-фосфатным остовом и четырьмя типами азотистых (пуриновых и пиримидиновых) оснований в качестве боковых групп (рис. 1). Существует только два небольших отличия цепи РНК от одиночной цепи ДНК:
1) пятиуглеродный сахар (пентоза) в РНК представлен рибозой, а в ДНК - его производным 2'-дезоксирибозой;
2) один из двух пиримидиновых нуклеотидов в РНК представлен уридиловым остатком (U) вместо его метилированного производного Т в ДНК.
Тот же вышеупомянутый принцип комплемен-тарности обеспечивает механизм репликации РНК на матрице ДНК. Разница лишь в том, что РНК полимеризуется только на одной из двух разошедшихся цепей двойной спирали ДНК (рис. 2). Разумеется, при синтезе РНК напротив А цепи ДНК становится уридиловый рибонуклеотид (U) вместо тимидилового дезоксирибонуклеотида (Т) при синтезе ДНК. Реплицирующаяся цепь РНК, таким образом, является точной копией противоположной цепи ДНК, с заменой Т на U. Процесс репликации сопровождается отделением цепи РНК от ДНК. В результате такой репликации РНК образуется как гибкий одноцепочечный полимер в отличие от жесткой двойной спирали ДНК.
Будучи копиями определенных функциональных отрезков цепи ДНК - генов, цепи РНК призваны служить матрицами для синтеза другого типа полимеров - полипептидных цепей белков. Так как белки состоят из двадцати разных сортов мономеров (аминокислот), а РНК - только из четырех сортов мономеров (нуклеотидов), детерминация аминокислотной последовательности полипептидной цепи нуклеотидной последовательностью РНК требует того, чтобы каждая аминокислота кодировалась комбинацией из нескольких - не менее трех - нуклеотидов. Именно триплетный код был сначала постулирован на основании теоретических соображений, а затем и доказан экспериментально. За РНК прочно закрепилась генетическая роль посредника между генами и белками: с одной стороны, РНК представлялась как совокупность копий генов, то есть копий отрезков ДНК, а с другой - как непосредственные матрицы, последовательности нуклеотидных триплетов которых декодируются в аминокислотные последовательности полипептидных цепей в процессе синтеза белков.
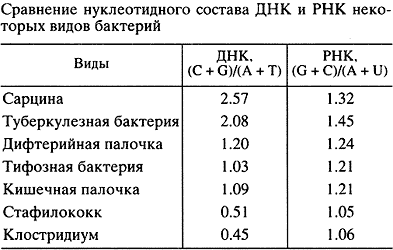
Исходя из этих представлений, в 1956 г. мной, тогда аспирантом А.Н. Белозерского в Институте биохимии АН СССР, была начата работа по экспериментальной проверке соответствия ДНК и РНК. Сначала мы показали, что нуклеотидный состав (соотношение четырех сортов нуклеотидов) ДНК может очень сильно различаться у разных видов организмов, в частности у бактерий разных таксономических групп. Далее мы исходили из того, что если РНК - копия ДНК, то нуклеотидный состав этих двух типов нуклеиновых кислот должен совпадать или, по крайней мере, быть сходным. Наши анализы дали совершенно неожиданный результат: при сильных вариациях состава ДНК соотношения нуклеотидов в тотальной РНК разных видов оказались удивительно консервативны (табл.) [14]. Вместе с тем статистический анализ этих данных показал, что имеется надежная положительная корреляция состава РНК с составом ДНК, хотя и при малой величине регрессии (рис. 3) [15]. Данные были интерпретированы так, что на фоне основной массы эволюционно консервативной, похожей у разных видов РНК существует относительно малая фракция видоспецифической РНК, копирующей ДНК.
В 1959 г. Ф. Крик так описал этот ранний период истории молекулярной биологии:
"Проблема кодирования прошла к настоящему времени три фазы. На первой - фазе блужданий - были сделаны различные предположения, но ни одно не было достаточно точным, чтобы подвергнуться опровержению. Вторая фаза - оптимистическая - была инициирована в 1954 году Гамовым, который был достаточно смелым, чтобы предложить довольно точный код. Это стимулировало целый ряд исследователей, стремившихся показать, что его предположения неверны, и тем самым они несколько подняли точность мышления в этой области. Третья фаза - фаза замешательства - была инициирована статьей Белозерского и Спирина в 1958 году... Данные, представленные там, показывали, что наши представления по многим важным аспектам были чересчур упрощенными" [16].
Таким образом, в работе 1956-1958 гг. нами были получены указания на два новых обстоятельства: во-первых, на наличие в клетках основной массы РНК, не являющейся посредником между генами и белками, то есть, по-видимому, негенетической РНК, и, во-вторых, на существование ДНК-подобной - генетической - РНК в виде сравнительно небольшой фракции, которой и могла быть приписана роль посредника между генами и белками.
Несколько ранее было установлено, что ДНК-подобная РНК образуется при заражении бактерий вирусом (бактериофагом): внедрение нового генетического материала - ДНК вируса - в клетку индуцировало синтез РНК, подобной по составу вирусной ДНК и, очевидно, определяющей синтез вирусных белков [17]. В нашей работе было впервые показано, что фракция ДНК-подобной РНК есть нормальный компонент обычных, не зараженных клеток, где она, по-видимому, выполняет функцию переноса генетической информации от своей ДНК, чтобы определять синтез своих белков. Позднее эта фракция РНК получила название messenger RNA [18], матричной РНК (мРНК), или информационной РНК.
Дальнейшие исследования мРНК в моей группе (с 1960 г. - лаборатории) в Институте биохимии АН СССР и переход от изучения микроорганизмов к высшим организмам привели еще к одному открытию. Оказалось, что в клетках высших организмов - животных и высших растений - мРНК в свободном виде не существует, она представлена в виде рибонуклеопротеидных частиц (мРНП-частиц), названных нами информосомами [19, 20].
Этот новый тип внутриклеточных частиц характеризовался рядом уникальных физико-химических свойств, и в частности, постоянным соотношением структурного белка и мРНК со значительным преобладанием белкового компонента. Роль нуклеопротеидной формы существования мРНК в клетках высших организмов в дальнейшем изучалась многими исследователями в связи с механизмами регуляции белкового синтеза на уровне трансляции. Эти работы дали ключ к пониманию ряда молекулярных механизмов оогенеза, сперматогенеза и раннего эмбриогенеза, клеточной дифференцировки и морфогенеза, эритропоэза и других процессов в жизнедеятельности многоклеточных существ, включая человека (см. обзоры [21, 22]).

