 2015-05-26
2015-05-26 3847
3847Рост и двигательная активность корня. Для выполнения своих функций — поглощения из почвы воды и минеральных веществ — корень должен прежде всего обладать способностью ориентироваться в пространстве, реагировать на градиенты жизненно необходимых факторов, продвигаться в соответствии с этим в почвенном слое и создавать максимальную поверхность для контакта с почвой.
При прорастании семян корень первоначально ориентируется в гравитационном поле, направляя свой рост к центру Земли (положительный геотропизм). Как показали опыты Ч. Дарвина и Н. Г. Холодного, чувствительность к направлению действия силы тяжести локализована в корневом чехлике. Действие гравитационного поля воспринимается статолитами (например, амилопластами), что оказывает влияние на транспорт АБК из кончика корня в зону растяжения (см. рис. 13.4). Одностороннее увеличение содержания АБК в зоне растяжения тормозит рост и приводит к целесообразным ростовым изгибам (см. 13.5.2). Избыток ИУК, поступающий из надземной части, также тормозит удлинение клеток в зоне растяжения, но способствует образованию боковых и придаточных корней.
Растущий дистальный конец корня очень чувствителен к механическим давлениям и проникает лишь в рыхлые участки почвы. При сильном механическом сопротивлении рост его в длину тормозится и одновременно в зоне растяжения появляется утолщение (из-за образования стрессового этилена). Это способствует расширению пор в почве и дальнейшему росту корня. Продвижение кончика корня между частицами почвы облегчает корневой чехлик, клетки которого постоянно се-кретируют во внешнюю среду слизистые вещества (полисахариды); кроме того, клетки ткани корневого чехлика и сами легко отделяются и разрушаются. Это не только способствует продвижению корня, но и защищает его от повреждений.
При недостатке воды именно она становится главным фактором, определяющим поведение корня. Вместо ориентации в гравитационном поле корень начинает реагировать на градиент влажности. Впервые это в простых и красивых опытах было показано Ю. Саксом в прошлом веке. Когда корни проростков, ориентирующихся первоначально в гравитационном поле, выходят снизу из косо подвешенного влажного блока, сделанного, например, из мха сфагнума (рис. 5.12), через некоторое время они изгибаются в сторону острого угла (где влажность больше) и начинают расти, приближаясь к блоку.
При недостатке воды резко увеличивается поверхность поглощающей зоны за счет интенсивного роста корневых волосков. Механизм этого явления еще не изучен. Зона корневых волосков (зона наибольшего поглощения) начинается после зоны растяжения. Вследствие этого зона поглощения перемещается в почве вслед за ростом дистальной, растущей части корня.
Таким образом, корни растений удлиняются, чтобы достичь новых участков почвы, и развивают большую поверхность соприкосновения с частицами почвенного поглощающего комплекса благодаря ветвлению, образованию все новых и новых поглощающих зон корней и корневых волосков.
Поглощение воды корнем и ее радиальный транспорт. Зона наиболее интенсивного поглощения воды совпадает с зоной развития корневых волосков. Основная функция корневых волосков заключается в увеличении всасывающей поверхности корня. Например, у 4-месячного растения ржи, выращенного в оптимальных условиях, насчитывалось в среднем 13800000 корней с общей площадью поверхности 232 м2, количество живых корневых волосков — 14 млрд. с площадью поверхности 399 м2. Суммарная площадь корней и корневых волосков составляла 631 м2 и они размещались в 0,05 м3 почвы.

Общая поверхность всей корневой системы в 130 раз превышала поверхность надземных частей того же самого растения. Даже если принять во внимание поверхность клеток мезофилла листьев, обращенную к межклетникам, то и в этом случае всасывающая поверхность корня будет в 22 раза больше, чем та площадь надземной части, где осуществляется транспирация.
Способность корневых волосков поглощать воду доказана экспериментально. Однако эпидермальные клетки, лишенные волосков, поглощают воду с такой же скоростью на единицу поверхности, как и клетки, несущие корневые волоски.
Выше зоны корневых волосков скорость всасывания воды снижается из-за опробковения клеток. Если начинается вторичный рост корня, то поглощение H2О обычно снижается еще сильнее. Однако и через опробковевшие участки корней вода частично транспортируется. У растений, обладающих микоризой, последняя также выполняет функцию дополнительной поглощающей поверхности, особенно в более старых частях корня.
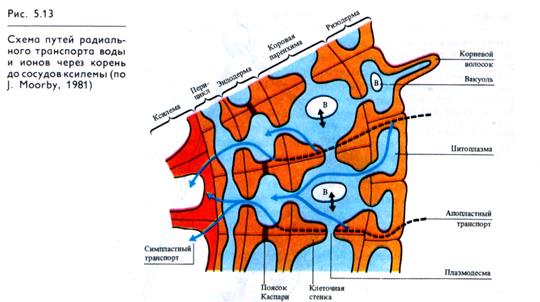
От поверхности корня через клетки коры, эндодерму и перицикл вода должна пройти до сосудов ксилемы. Через клетки коры возможны два пути транспорта воды и минеральных веществ (рис. 5.13): через цитоплазму по плазмодесмам (сим-пластный транспорт) и по клеточным стенкам (апопластный транспорт). Вода поступает в цитоплазму клеток ризодермы и паренхимных клеток корня по законам осмоса.
Поскольку сопротивление клеточных стенок для воды значительно ниже, чем у цитоплазмы, более быстрый радиальный транспорт воды осуществляется через корень по апопла-сту. Однако на уровне эндодермы этот тип транспорта становится невозможным из-за непроницаемых для воды поясков Каспари. Следовательно, вода может преодолеть эндодерму только пройдя через мембраны и цитоплазму этих клеток. Регуляция подачи воды на уровне эндодермы осуществляется, с одной стороны, сменой быстрого апопластного транспорта на медленный симпластный, а с другой — тем, что диаметр стели, куда должна подаваться вода через эндодерму, в 5 —6 раз меньше диаметра поверхности коры и всасывающей поверхности корня. Следует отметить, что непроницаемость клеточных стенок эндодермы для воды не является абсолютной. Так, в растущих зонах корня, где пояски Каспари клеток эндодермы сформированы не полностью, смены типов транспорта, по-видимому, не происходит. Кроме того, в участках корня, где закладываются боковые корни, эндодерма прерывается. Однако в целом массовый ток воды через эндодерму по апопласту резко ограничен.
 Внутри проводящего цилиндра движение воды от эндодермы до сосудов ксилемы встречает небольшое сопротивление и, возможно, осуществляется по клеточным стенкам.
Внутри проводящего цилиндра движение воды от эндодермы до сосудов ксилемы встречает небольшое сопротивление и, возможно, осуществляется по клеточным стенкам.
Механизмы корневого давления. В сосуды ксилемы вода поступает, как и на первых этапах поглощения, благодаря осмотическому механизму. Существование таких механизмов в транспорте воды доказывается тем, что если во внешней среде создать осмотическую концентрацию, равную внутриклеточной, то вода в клетки поступать не будет; если осмотическая концентрация в среде выше, вода выходит из клеток, если ниже — идет ее поглощение. Осмотически активными веществами в сосудах и их клеточных стенках служат минеральные вещества и метаболиты, выделяемые активными ионными насосами, функционирующими в плазмалемме паренхимных клеток, окружающих сосуды (см. 8.1.2). Накопление этих осмотически активных веществ в сосудах создает сосущую силу, способствующую осмотическому транспорту воды в ксилему.
Сосущая сила сосудов может оказаться выше, чем у окружающих их живых клеток, не только из-за повышающейся концентрации ксилемного сока, но также из-за отсутствия противодавления со стороны клеточных стенок, которые в сосудах лигнифицированы и, следовательно, не эластичны.
Таким образом, в результате активной работы ионных насосов в корне и осмотическому (пассивному) поступлению воды в сосуды ксилемы в сосудах развивается гидростатическое давление, получившее название корневого давления. Оно обеспечивает поднятие ксилемного раствора по сосудам ксилемы из корня в надземные части. Показано, что у растений, обитающих в холодных и плохо аэрируемых почвах, а также в результате действия ядов и ингибиторов корневое давление снижено.
Механизм поднятия воды по растению вследствие развивающегося корневого давления называют нижним концевым двигателем.
«Плач» растений. Гуттация. Примером работы нижнего концевого двигателя служит так называемый «плач» растений. Весной у деревьев с еще нераспустившимися листьями можно наблюдать интенсивный ксилемный ток жидкости снизу вверх через надрезы ствола и даже верхних веток кроны (раненое дерево «плачет»). Явление «плача» свидетельствует о значительном корневом давлении, которое в этот период у основания ствола достигает не менее 1,013 МПа (10 атм). У вегетирующих растений при удалении стебля с листьями из оставшегося связанного с корнем пенька довольно долго выделяется ксилемный сок, или пасока. Метод сбора пасоки служит одним из экспериментальных приемов по изучению функционирования корневых систем. Корневое давление можно измерить, поместив на пенек манометр (рис. 5.14, А).
Другим примером работы нижнего концевого двигателя является гуттация. При высокой влажности воздуха в результате деятельности нижнего концевого двигателя на концах и зубчиках листьев выделяется капельно-жидкая влага — растение гуттирует (рис. 5.14, Б). Гуттационное выделение влаги листьями особенно характерно для тропических растений, приспособленных к жизни в условиях повышенной влажности, при которых транспирация затруднена. В этих условиях подъ ем воды практически осуществляется только вследствие корневого давления.

 Под такими деревьями в тропическом лесу как будто постоянно идет дождь. Функцию выделения жидкости из тканей листьев выполняют специальные образования — гидатоды, локализованные в зубчиках листьев. Ксилемный сок из подходящих к гидатоде сосудов ксилемы под действием корневого давления поступает в мелкоклеточную паренхимную ткань гидатоды — эпитему, проходит через нее, обедняясь особенно азотсодержащими веществами, и через субэпидермаль-ную полость и водяное устьице выделяется наружу. В гутта-ционной жидкости обнаружены минеральные и органические соединения.
Под такими деревьями в тропическом лесу как будто постоянно идет дождь. Функцию выделения жидкости из тканей листьев выполняют специальные образования — гидатоды, локализованные в зубчиках листьев. Ксилемный сок из подходящих к гидатоде сосудов ксилемы под действием корневого давления поступает в мелкоклеточную паренхимную ткань гидатоды — эпитему, проходит через нее, обедняясь особенно азотсодержащими веществами, и через субэпидермаль-ную полость и водяное устьице выделяется наружу. В гутта-ционной жидкости обнаружены минеральные и органические соединения.
Трапспирация — это физиологический процесс испарения воды растением. Основным органом транспирации является лист.
Из-за необходимости максимального контакта с воздушной средой растение имеет очень большую листовую поверхность. Увеличение поверхности облегчает поглощение СО2, улавливание света и это же создает огромную поверхность испарения. Вода испаряется с поверхности листьев и через устьица (см. рис. 13.10). В результате потери воды клетками листьев в них снижается водный потенциал, т. е. возрастает сосущая сила. Это приводит к усилению поглощения клетками листа воды из ксилемы жилок и передвижению воды по ксилеме из корней в листья.
Таким образом, верхний концевой двигатель, обеспечивающий передвижение воды вверх по растению, создается и поддерживается высокой сосущей силой транспирирующих клеток листовой паренхимы. Поэтому сила верхнего концевого двигателя будет тем больше, чем активнее транспирация. Верхний концевой двигатель может работать при полном отключении нижнего концевого двигателя, причем для его работы используется не метаболическая энергия, а энергия внешней среды — температура и движение воздуха.
 Лист как орган транспирации. Из стебля вода движется в лист через черешок или листовое влагалище, а затем по листовым жилкам. Количество проводящих элементов в каждой жилке последовательно уменьшается по мере их ветвления.
Лист как орган транспирации. Из стебля вода движется в лист через черешок или листовое влагалище, а затем по листовым жилкам. Количество проводящих элементов в каждой жилке последовательно уменьшается по мере их ветвления.
Самые мелкие жилки состоят из единичных трахеид. Система их распределения настолько эффективна, что редко клетки листа отделены от сосудистых элементов более чем двумя другими клетками. В листьях некоторых растений, особенно с С4-путем фотосинтеза, сосудистые пучки окружены одним слоем компактно расположенных паренхимных клеток, которые образуют обкладку пучка и одновременно выполняют функцию механической ткани. Обкладка тянется до самого окончания каждого сосудистого пучка. Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой, а часто и восковым налетом. Эпидермис вместе с кутикулой образует эффективный барьер на пути движения воды. На поверхности эпидермиса листьев часто развиты волоски, чешуйки, которые также влияют на водный режим листа, так как сильно снижают скорость движения воздуха над его поверхностью и рассеивают свет, что снижает потери воды за счет транспирации.
Как отмечалось (см. рис. 3.17), мезофилл листа состоит из палисадной и губчатой паренхимы с большими межклетниками. Хотя палисадная ткань плотнее, чем губчатая, поверхность ее клеток также соприкасается с воздухом в межклетниках губчатой паренхимы. Площадь внутренней поверхности листа обычно на порядок больше площади его наружной поверхности. Вода в межклетниках испаряется со всех открытых участков клеток мезофилла.
Транспирация слагается из двух процессов: а) передвижения воды из листовых жилок в поверхностные слои стенок клеток мезофилла; б) испарения воды из клеточных стенок в межклетные пространства и подустьичные полости с последующей диффузией в окружающую атмосферу через устьица (устьичнан транспирация) или испарения воды из клеточных стенок эпидермиса в атмосферу путем кутикулярной транспирации.
Жидкая вода транспортируется к испаряющим поверхностям преимущественно по клеточным стенкам. Как и в корне, это обусловлено тем, что в клеточных стенках вода встречает более слабое сопротивление, чем на пути от клетки к клетке через протопласты и вакуоли. В межклетниках воздух насыщен водой наполовину, а водный потенциал уравновешен с водным потенциалом окружающих клеток, где он редко бывает ниже 2 МПа. Водный потенциал атмосферного воздуха тем ниже (более отрицателен), чем меньше его относительная влажность. Молекулы воды покидают растение, перемещаясь (как и внутри растения) в направлении более низкого водного потенциала, т. е. из тканей наружу через устьица.
Устьичная транспирация. Устьица играют важнейшую роль в газообмене между листом и атмосферой, это основной проводящий путь для водяного пара, СО2 и О2. Устьица могут находиться на обеих сторонах листа, но есть виды, у которых устьица располагаются только на нижней стороне листа. Даже на одном растении у затененных листьев устьиц меньше, чем у постоянно хорошо освещаемых («световых») листьев. В среднем число устьиц колеблется от 50 до 500 на  1 мм2. Транспирация с поверхности листа через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий и мала по отношению к площади всего листа (0,5 — 2,0%), испарение воды через многочисленные устьица идет очень интенсивно.
1 мм2. Транспирация с поверхности листа через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий и мала по отношению к площади всего листа (0,5 — 2,0%), испарение воды через многочисленные устьица идет очень интенсивно.
Кутикулярная транспирация. При открытых устьицах потери водяного пара через кутикулу листа обычно незначительны по сравнению с общей транспирацией. Но если устьица закрыты, как, например, во время засухи, кутикуляр-ная транспирация приобретает важное значение в водном режиме растений многих видов. Интенсивность кутикулярной транспирации сильно варьирует у разных видов: от совершенно незначительных потерь до 50% от общей транспирации. У молодых листьев с тонкой кутикулой кутикулярная транспирация составляет около половины всей транспирации. У зрелых листьев с более мощной кутикулой кутикулярная транспирация составляет лишь '/ю общей транспирации. В стареющих листьях кутикулярное испарение воды может вновь возрастать из-за разрушения и растрескивания кутикулы. Кутикулярная транспирация регулируется главным образом толщиной слоя кутикулы. Виды магнолий и хвойных, обладающие толстыми слоями кутина в кутикуле, теряют очень мало воды через эпидермис листа. И наоборот, виды с тонким слоем кутина продолжают терять воду и после того, как устьица закроются, и поэтому гораздо сильнее страдают от засухи.
Некоторое количество воды выделяется в результате транспирации почек, поскольку почечные чешуи полностью не предохраняют их от потери влаги. Репродуктивные органы также теряют воду и в некоторых случаях эти потери могут быть очень значительными: например, корзинки подсолнечника, коробочки мака и плоды перца транспирируют сильнее, чем листья данных растений в тех же условиях. Кроме того, вода испаряется с поверхности ветвей и стволов древесных растений через чечевички и окружающие их слои пробки (перидермалъная транспирация). Хотя общее количество воды, испаряемой через чечевички, значительно меньше того, которое теряется через листья, фактически интенсивность транспирации на единицу испаряющей поверхности нередко мало различается в обоих случаях. Вследствие транспирации ветвей в зимнее время часто возникает водный дефицит и растения гибнут в результате обезвоживания.
Регуляция устьичной транспирации. Открывание устьиц регулируется несколькими взаимодействующими механизмами.
Движущей силой, вызывающей изменение ширины устьичной щели (отверстости устьиц), является изменение тургора замыкающих (иногда и прилегающих к устьицам) клеток. По мере того как замыкающая клетка устьица осмотически поглощает воду, более тонкая и эластичная часть ее клеточной стенки, удаленная от щели, растягивается. Поскольку более
 толстый и менее эластичный участок стенки, окаймляющий щель, растягивается слабее, замыкающие клетки принимают полукруглую форму, в результате чего устьица раскрываются (см. рис. 13.10). Факторы внешней и внутренней среды прямо и косвенно воздействуют на устьичный аппарат, вызывая в замыкающих клетках изменения, которые в свою очередь приводят к изменению тургора. Из внешних факторов на движения устьиц больше всего влияют влажность воздуха и условия водоснабжения, свет и температура, а из внутренних — парциальное давление СО2 в системе межклетников, состояние гидратации растения, ионный баланс и фитогормоны, из которых цитокинин способствует открыванию устьиц, а абсцизовая кислота — закрыванию. На состояние устьиц влияют возраст листьев и фазы развития растения, а также эндогенные суточные ритмы.
толстый и менее эластичный участок стенки, окаймляющий щель, растягивается слабее, замыкающие клетки принимают полукруглую форму, в результате чего устьица раскрываются (см. рис. 13.10). Факторы внешней и внутренней среды прямо и косвенно воздействуют на устьичный аппарат, вызывая в замыкающих клетках изменения, которые в свою очередь приводят к изменению тургора. Из внешних факторов на движения устьиц больше всего влияют влажность воздуха и условия водоснабжения, свет и температура, а из внутренних — парциальное давление СО2 в системе межклетников, состояние гидратации растения, ионный баланс и фитогормоны, из которых цитокинин способствует открыванию устьиц, а абсцизовая кислота — закрыванию. На состояние устьиц влияют возраст листьев и фазы развития растения, а также эндогенные суточные ритмы.
Сильнейшее влияние на движения устьиц оказывает степень обеспеченности клетки водой. Различают гидропассивную и гидроактивную устьичные реакции. Активными называют движения, зависящие от изменений в самих замыкающих клетках, пассивными — движения, определяемые изменениями в клетках, окружающих устьичные. Гидропассивное закрывание устьиц связано со сдавливающим действием соседних клеток эпидермиса (и хлоренхимы) в условиях их полного тургора (при высокой насыщенности водой). Гидропассивное открывание устьиц может произойти при ослаблении этого сдавливания в условиях слабого дефицита воды.
Гидроактивное закрывание устьиц произойдет, как только транспирация превысит поглощение воды корнями и снижение тургора в замыкающих клетках достигнет критического уровня. Этот уровень имеет разную величину в зависимости от вида растения, возраста листьев и степени приспособленности к окружающей среде. Реакция закрывания устьиц по мере развития водного дефицита в тканях обусловлена увеличением концентрации фитогормона — абсцизовой кислоты в клетках листа (см. 2.5.4). Абсцизовая кислота подавляет деятельность Н +-насосов на плазмалемме замыкающих клеток, вследствие чего снижается их тургор и устьица закрываются (более подробно см. 13.6.1). Для усиления синтеза АБК достаточно снижения водного потенциала листа на 0,2 МПа, что еще не выражается в видимом завядании, но приводит к закрыванию устьиц. При нанесении АБК на основание листа устьица закрываются через 3 — 9 мин.
В отличие от других клеток эпидермиса замыкающие клетки устьиц содержат хлоропласты. На свету при хорошем водоснабжении устьица открываются тем шире, чем больше интенсивность освещения (механизм фотоактивного открывания), причем действующим фактором является синий свет. Фотосинтез в замыкающих клетках также участвует в регуляции устьичных движений. Усиление синтеза углеводов в замыкающих клетках увеличивает их сосущую силу и вызывает поглощение воды, способствуя этим открыванию устьиц. Обратимые превращения крахмала в сахар существенны в измене нии сосущей силы и тургора в замыкающих клетках.
Открывание устьиц в утренние часы регулируется главным образом светом. Как правило, после полудня по мере усиления напряженности водного дефицита устьица закрываются.
Состояние устьиц зависит и от СО2. Если концентрация СО2 в подустьичной полости падает ниже 0,03%, тургор замыкающих клеток увеличивается и устьица открываются. Частично с этим связано открывание устьиц с восходом солнца: усиление фотосинтеза снижает концентрацию СО2 в межклетниках. Закрывание устьиц можно вызвать повышением концентрации СО2 в воздухе. Это происходит в межклетниках листа ночью, когда в результате отсутствия фотосинтеза и продолжающегося дыхания уровень СО2 в тканях повышается. Такого рода регуляция устьичных движений СО2 позволяет понять, почему устьица закрыты ночью и открываются с восходом солнца.
Решающее влияние концентрации СО2 на степень открытости устьиц обнаруживается у суккулентов, обладающих специфическим суточным ритмом обмена органических кислот. Эти растения открывают устьица ночью, когда парциальное давление СО2 в межклетниках их листьев снижается вследствие интенсивного образования малата, а закрывают устьица, когда при декарбоксилировании малата днем высвобождается СО2, который накапливается в межклетниках перед дальнейшим использованием.
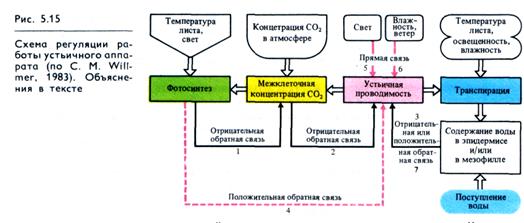
Таким образом, в регуляции функционирования устьиц (рис. 5.15) взаимодействуют прямые и обратные связи. Одна из них (2) предотвращает недостаток СО2, который может быть вызван фотосинтезом. Когда межклеточная концентрация СО2 снижается до уровня, недостаточного для фотосинтеза, это служит сигналом обратной связи и устьица открываются для обмена СО2 с внешней средой. Другой тип связи (3) реагирует на содержание воды в листьях: при снижении содержания воды в тканях устьица закрываются. В результате деятельности этих двух типов обратных связей обычно наблюдаются небольшие осцилляции отверстости устьиц. Однако одни обратные связи по СО2 (1,2) и Н2О (3) не оптимизируют газовый обмен листа.
Оптимизация достигается также деятельностью прямой связи — прямого действия света (5), влажности и ветра (6) и положительной обратной связи фотосинтеза (4) и водного режима эпидермальных клеток и (или) клеток мезофилла (7). Важным механизмом функционирования петли отрицательной обратной связи (3) является синтез АБК. Ряд продуктов фотосинтеза в клетках мезофилла (и в устьичных клетках) может контролировать движения устьиц по механизму положительной обратной связи (4).
Суточные колебания транспирации. Периодичность суточного хода транспирации наблюдается у всех растений, но кривые, отражающие фактическую транспирацию, сильно отличаются у разных видов и в неодинаковых погодных условиях.
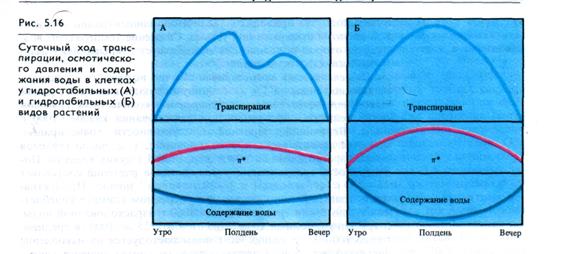
У деревьев, теневыносливых растений, многих злаков и т. д. (гидростабильные виды) с совершенной регуляцией устьичной транспирации (рис. 5.16, А) испарение воды достигает максимума до установления максимума дневной температуры. В полуденные часы транспирация падает и вновь может увеличиваться в предвечерние часы при снижении температуры воздуха. Такой ход транспирации приводит к незначительным суточным изменениям осмотического давления и содержания воды в листьях. У видов, способных переносить резкие изменения содержания воды в клетках в течение дня (гидролабильные виды), наблюдается одновершинный суточный ход транспирации с максимумом в полуденные часы (рис. 5.16, Б). В обоих случаях ночью транспирация минимальна.
Колебания интенсивности транспирации отражают изменения степени открытия устьиц в течение суток. Закрывание устьиц в полдень может быть связано как с увеличением уровня СО2 в листьях при повышении температуры воздуха (из-за усиления дыхания и фотодыхания), так и с возможным водным дефицитом, возникающим в тканях при высокой температуре, низкой влажности воздуха и особенно в ветреную погоду. Как  отмечалось, это приводит к увеличению концентрации абсцизо-вой кислоты и закрыванию устьиц. Снижение температуры воздуха во второй половине дня способствует открыванию устьиц и усилению фотосинтеза.
отмечалось, это приводит к увеличению концентрации абсцизо-вой кислоты и закрыванию устьиц. Снижение температуры воздуха во второй половине дня способствует открыванию устьиц и усилению фотосинтеза.
Интенсивность транспирации обычно выражают в граммах испаренной воды за 1 ч на единицу площади или на 1 г сухой массы; продуктивность транспирации — количеством граммов сухих веществ, образуемых при расходовании каждых 1000 г воды. Величиной, обратной продуктивности транспирации, является транспирационный коэффициент, т. е. число граммов воды, израсходованной при накоплении 1 г сухих веществ. Интенсивность транспирации у большинства растений составляет 15 — 250 г • м-2 ч-1 днем и 1—20 г • м-2. ч-1 ночью. Продуктивность транспирации у растений в умеренном климате колеблется от 1 до 8 г (в среднем 3 г) на 1000 г израсходованной воды, а транспирационный коэффициент — от 125 до 1000 (в среднем, около 300 г, т. е. около 300 г воды расходуется на накопление 1 г сухих веществ). Следовательно, на синтез веществ своего тела растение использует лишь 0,2% пропускаемой воды, остальные 99,8% тратятся на испарение.
Восходящий поток воды движется по растению главным образом по ксилеме (см. также 8.1). Ксилема представляет собой распределительную систему, снабжающую все ткани и органы растения водой. Как отмечалось, элементы ксилемы, формирующиеся из прокамбиальных клеток корня и стебля, в растущих растяжением кончиках корней содержат цитоплазму. Зрелые сосуды и трахеиды с одревесневшими клеточными стенками лишены цитоплазмы и выполняют водопроводящую функцию. Помимо восходящего тока воды по ксилеме существует ток воды по клеточным стенкам живых клеток от корня до листа, также поддерживаемый градиентом водного потенциала, создаваемым транспирацией. Доля этого потока воды составляет 1 — 10% общего потока.
Характеристики проводящего пути в стебле исследовались с помощью наблюдений за движением меченой воды, добавленной в среду, где находились корни. Вода в ксилеме быстро замещается меченой водой. Одновременно с прохождением меченой воды через сосудистую ткань последовательно замещается вода и в остальных тканях стебля, т. е. происходит обмен молекулами воды между движущимся потоком и всеми клетками.
 1\ Теория сцепления. Движущей силой восходящего тока воды в проводящих элементах ксилемы является градиент водного потенциала через растение от почвы до атмосферы. Он поддерживается двумя основными компонентами: 1) градиентом осмотического потенциала в клетках корня (от почвы до сосудов ксилемы), создаваемым активным транспортом ионов в живых клетках корня, включая молодые живые элементы ксилемы, и 2) транспирацией. Поддержание первого градиента требует затрат метаболической энергии; на транспирацию используется энергия солнечной радиации. Градиент осмотического потенциала обеспечивает поглощение воды корнем. Транспирация
1\ Теория сцепления. Движущей силой восходящего тока воды в проводящих элементах ксилемы является градиент водного потенциала через растение от почвы до атмосферы. Он поддерживается двумя основными компонентами: 1) градиентом осмотического потенциала в клетках корня (от почвы до сосудов ксилемы), создаваемым активным транспортом ионов в живых клетках корня, включая молодые живые элементы ксилемы, и 2) транспирацией. Поддержание первого градиента требует затрат метаболической энергии; на транспирацию используется энергия солнечной радиации. Градиент осмотического потенциала обеспечивает поглощение воды корнем. Транспирация
служит главной движущей силой восходящего тока воды, так как создает большой отрицательный градиент гидростатического давления в ксилеме, который реализуется в возникающем натяжении воды в сосудах ксилемы.
 Согласно теории сцепления (когезионной теории), предложенной в конце XIX в., вода в капиллярных трубках сосудов ксилемы поднимается вверх в ответ на присасывающее действие транспирации вследствие действия сил сцепления (когезии) молекул воды друг с другом и действия сил прилипания (адгезии) столба воды к гидрофильным стенкам сосудов. Обе силы препятствуют также образованию полостей у стенок сосудов, заполненных воздухом (или парами воды) и способных закупорить сосуд. При закупорке (эмболии) сосудов пузырьками воздуха всегда остается достаточное количество интактных нитей воды в других сосудах, чтобы обеспечить ток вверх. Кроме того, по-видимому, существуют механизмы восстановления непрерывности нарушенных эмболией водных нитей. Было показано, что сопротивление воды разрыву при 20 °С составляет 30 МПа. Этого давления достаточно (с учетом потерь на трение), чтобы обеспечить подъем воды на высоту 120—130 м.
Согласно теории сцепления (когезионной теории), предложенной в конце XIX в., вода в капиллярных трубках сосудов ксилемы поднимается вверх в ответ на присасывающее действие транспирации вследствие действия сил сцепления (когезии) молекул воды друг с другом и действия сил прилипания (адгезии) столба воды к гидрофильным стенкам сосудов. Обе силы препятствуют также образованию полостей у стенок сосудов, заполненных воздухом (или парами воды) и способных закупорить сосуд. При закупорке (эмболии) сосудов пузырьками воздуха всегда остается достаточное количество интактных нитей воды в других сосудах, чтобы обеспечить ток вверх. Кроме того, по-видимому, существуют механизмы восстановления непрерывности нарушенных эмболией водных нитей. Было показано, что сопротивление воды разрыву при 20 °С составляет 30 МПа. Этого давления достаточно (с учетом потерь на трение), чтобы обеспечить подъем воды на высоту 120—130 м.
Факт существования когезионного натяжения воды в транс-пирирующих растениях подтверждается многими наблюдениями. Опыты показали, что облиственный побег, срезанный под водой и герметически присоединенный к верхней части ртутного манометра, вызывает подъем ртути на высоту, превышающую барометрическую. Другим доказательством существования натяжения может служить сокращение диаметра древесного ствола, наблюдаемое в условиях интенсивной транспирации, так как натяжение нитей воды в сосудах приводит к уменьшению их диаметров.
Абсолютная скорость передвижения воды по ксилеме невелика. Для лиственных пород она составляет в среднем 20 см3 • ч-1 на 1 см2 поперечного сечения древесины, а для хвойных — 5 см3 • ч-1 (для сравнения: скорость движения крови по артериям равна 40-50 см3 • с-1). Однако такая скорость снижает сопротивление движению воды, возникающее в капиллярах сосудов древесины, и сосущая сила клеток листьев оказывается достаточной как для удержания «на весу» всей массы воды в древесине, так и для транспорта ее вверх.

