 2015-05-26
2015-05-26 1775
1775 Водообмен между ксилемой и флоэмой в целом растении. Вода и минеральные вещества доставляются к каждой клетке надземной части растения благодаря восходящему току воды по ксилеме (называемому иногда восходящим массовым током). Большая часть воды тока испаряется в атмосферу в процессе транспирации, а меньшая (около 0,2%) используется: 1) на метаболические реакции, входя в состав синтезируемых растением органических соединений; 2) на поддержание тургора клеток; 3) в транспорте органических веществ по флоэме от листьев к корням. Направленный вниз флоэмный ток растворов формируется в клетках мезофилла листьев (подробнее см. 8.2), где часть воды, доставленной ксилемным током, из клеточных стенок мезофилла переходит во флоэмные окончания.
Водообмен между ксилемой и флоэмой в целом растении. Вода и минеральные вещества доставляются к каждой клетке надземной части растения благодаря восходящему току воды по ксилеме (называемому иногда восходящим массовым током). Большая часть воды тока испаряется в атмосферу в процессе транспирации, а меньшая (около 0,2%) используется: 1) на метаболические реакции, входя в состав синтезируемых растением органических соединений; 2) на поддержание тургора клеток; 3) в транспорте органических веществ по флоэме от листьев к корням. Направленный вниз флоэмный ток растворов формируется в клетках мезофилла листьев (подробнее см. 8.2), где часть воды, доставленной ксилемным током, из клеточных стенок мезофилла переходит во флоэмные окончания.
Вода из клеток листа и непосредственно из сосудов ксилемы поступает во флоэму по осмотическому градиенту, возникающему вследствие накопления в клетках флоэмы Сахаров и других органических соединений, образуемых в процессе фотосинтеза.
Нисходящий флоэмный ток доставляет органические соединения тканям корня, где они используются в метаболизме. В корне в окончаниях проводящих пучков элементы флоэмы, как и в листе, располагаются вблизи элементов ксилемы и вода вновь по осмотическому градиенту поступает в ксилему и движется вверх с восходящим током. Таким образом происходит обмен воды в проводящей системе корня и листьев.
Постоянный ток воды от корневой системы к надземным частям, во-первых, служит средством транспортировки и накопления в надземных органах минеральных веществ и химических соединений, поступающих из корней. Используя энергию солнца (нагрев) и движение воздуха, растение транспирирует воду, создавая таким образом условия для переноса с током воды питательных веществ. Транспирация и усвоение углерода связаны друг с другом. Чтобы получить СО2 (открывание устьиц), растение вынуждено отдавать воду, а уменьшение потери воды (закрывание устьиц) снижает и приток СО2. В сельском и лесном хозяйстве для получения максимальных урожаев важно знать соотношение между продукцией фотосинтеза и расходованием воды у выращиваемых растений с целью их регуляции.
Во-вторых, восходящий ток воды необходим для нормального водоснабжения всех клеток и поддержания тургора. При недостатке водоснабжения в структуре и функциональной активности клеток наблюдаются различные нарушения (см. 14.3). Поэтому для получения устойчивых урожаев в засушливых районах необходимо развивать орошаемое земледелие. В условиях высокой сухости почвы большую роль в поддержании водного режима растений может играть их опрыскивание (создание искусственной росы).
В-третьих, транспирация может быть способом защиты от перегрева. Однако этот вопрос до конца не разрешен. Известно, что растения превосходно рассеивают теплоту и не перегреваются во влажных теплицах при незначительной транспирации.

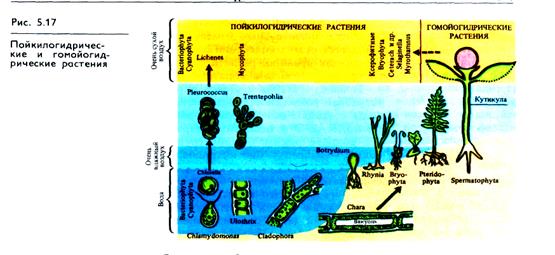
Разнообразие типов водного обмена у растений (рис. 5.17) можно представить в виде следующей схемы:
Растения, обитающие в воде (гидратофиты), регулируют постоянство состава внутренней среды с помощью механизмов защиты от избыточного поступления воды (см. 13.7), которую они поглощают всей поверхностью. Первичными гидратофита-ми являются водоросли. Водные цветковые растения (вторичные гидратофиты, происходящие от наземных форм) совмещают черты настоящих гидратофитов с чертами, свойственными высшим наземным растениям.
 Наземным растениям, непрерывно испаряющим воду, необходим уравновешенный водный баланс, поэтому механизмы его регуляции направлены на защиту от избыточной потери воды. Однако они неодинаковы у растений разных экологических групп. По способности приспосабливать водный обмен к колебаниям водоснабжения различают две группы растений (Г. Вальтер): пойкилогидрические и гомойогидрические.
Наземным растениям, непрерывно испаряющим воду, необходим уравновешенный водный баланс, поэтому механизмы его регуляции направлены на защиту от избыточной потери воды. Однако они неодинаковы у растений разных экологических групп. По способности приспосабливать водный обмен к колебаниям водоснабжения различают две группы растений (Г. Вальтер): пойкилогидрические и гомойогидрические.
Пойкилогидрические растения (бактерии, синезе-леные водоросли, низшие зеленые водоросли порядка Protococ-cales, грибы, лишайники и др.) приспособились переносить значительный недостаток воды без потери жизнеспособности. При этом у них снижается интенсивность обмена веществ, клетки равномерно сжимаются. Увеличение количества воды в среде приводит к возобновлению активного метаболизма в клетках пойкилогидрических растений. По характеру изменения показателей водного режима (интенсивность транспирации, осмотическое давление, содержание воды) в течение суток они относятся к гидролабильным растениям (см. рис. 5.16, Б), так как у них значительно колеблется содержание воды и испарение. К этой группе относятся лишь немногие покрытосеменные (например, злаки сухих степей), однако их пыльцевые зерна и зародыши (семена) пойкилогидричны.
Гомойогидрические растения (наземные папоротни-кообразные, голосеменные, цветковые) составляют большинство обитателей суши. Они обладают тонкими механизмами регуляции устьичной и кутикулярной транспирации, а также корневой системой, обеспечивающей поставку воды. Поэтому даже при значительных изменениях влажности среды не наблюдается резких колебаний содержания воды в клетках, у которых, как правило, развита вакуолярная система. При этом клетки не способны к обратимому высыханию.
Точная регуляция поставок и трат воды этими растениями устраняет возможность значительных колебаний содержания воды в тканях, осмотического давления и транспирации (см. рис. 5.16, А). Эти показатели характеризуют гидростабильный тип водного режима данной группы растений. Стабилизации водного режима у многих видов способствуют запасы воды в корнях, стеблях, запасающих органах и т. д. Гомойогидрические растения делятся на три экологические группы:
1. Гигрофиты (тонколистные папоротники, некоторые фиалки, калужница и др.), произрастающие в условиях повышенной влажности и (или) недостаточной освещенности. Теневыносливые гигрофиты, с почти всегда открытыми устьицами, имеют гидатоды, через которые выделяют избыток воды в ка-пельно-жидком состоянии. Гигрофиты плохо переносят почвенную и воздушную засуху.
2. Ксерофиты (молочай, алоэ, кактусы, полынь, ковыль и др.) преобладают в местностях с жарким и сухим климатом и хорошо приспособлены к перенесению атмосферной и почвенной засухи (см. 14.3).
3. Мезофиты (лиственные деревья, лесные и луговые травы, большинство культурных растений и т. д.) по способности регулировать водный режим занимают промежуточное положение между гидрофитами и ксерофитами (см. 14.3).
Развитие и выживание растений в любых условиях гораздо сильнее зависит от доступности воды, чем от какого-либо иного фактора внешней среды. Примерно '/з поверхности суши занимают области, где обнаруживается дефицит влаги; половина этой площади (около 12% поверхности Земли) является крайне засушливой. Районы с избытком осадков занимают менее 9% поверхности суши. Если годовые осадки превышают годовое испарение, то говорят о гумидной зоне, в обратном случае — об аридной. Многие сельскохозяйственные районы расположены в аридных зонах, где земледелие возможно только благодаря искусственному орошению.
На протяжении многих лет считалось доказанным, что в пересыхающей почве вода доступна растениям до тех пор, пока содержание влаги в ней не достигнет коэффициента устойчивого завядания, когда в почве остается недоступная растению вода. Согласно этой точке зрения физиологические процессы, рост и развитие растений на почве, подвергающейся иссушению, протекают нормально до достижения коэффициента завядания. Однако накоплено много данных, показывающих, что на обмен веществ, а следовательно, на рост и развитие растений влияет даже слабый водный дефицит. Такой внутренний водный дефицит возникает в тканях задолго до того, как содержание влаги в почве приблизится к уровню коэффициента завядания.
 Растения, перенесшие только однократную сильную кратковременную засуху, так и не возвращаются к нормальному обмену веществ. Внутренний водный баланс растения зависит от комплекса факторов, связанных а) с самим растением (засухоустойчивость, глубина проникновения и ветвление корней, фаза развития); б) с количеством растений на данной площади; в) с климатическими факторами (потери воды на испарение и транспирацию, температура и влажность воздуха, туман, ветер и свет, количество осадков и т. д.); г) с почвенными факторами (количество воды в почве, осмотическое давление почвенного раствора, структура и влагоемкость почвы и др.).
Растения, перенесшие только однократную сильную кратковременную засуху, так и не возвращаются к нормальному обмену веществ. Внутренний водный баланс растения зависит от комплекса факторов, связанных а) с самим растением (засухоустойчивость, глубина проникновения и ветвление корней, фаза развития); б) с количеством растений на данной площади; в) с климатическими факторами (потери воды на испарение и транспирацию, температура и влажность воздуха, туман, ветер и свет, количество осадков и т. д.); г) с почвенными факторами (количество воды в почве, осмотическое давление почвенного раствора, структура и влагоемкость почвы и др.).
Дефицит влаги в растениях действует на такие процессы, как поглощение воды, корневое давление, прорастание семян, устьичные движения, транспирация, фотосинтез, дыхание, ферментативная активность растений, рост и развитие, соотношение минеральных веществ и др. Изменяя обмен веществ, недостаток воды влияет на продуктивность, вкус плодов, плотность древесины, длину и прочность волокна у хлопчатника и т. д.
Степень оводненности, необходимая для начала прорастания семян, сильно варьирует у разных видов. Между скоростью прорастания семян и скоростью поглощения воды существует определенная корреляция. На скорость прорастания влияют также свойства семян и водоудерживающая способность почвы.
Влияние водного дефицита на метаболические процессы в значительной мере зависит от длительности его действия. При устойчивом завядании растений увеличивается скорость распада РНК, белков и одновременно возрастает количество небелковых азотсодержащих соединений и отток их в стебель и колос (у злаков). В результате в условиях засухи содержание белков в листьях относительно уменьшается, а в семенах — увеличивается.
Влияние водного дефицита на углеводный обмен выражается вначале в снижении содержания моно- и дисахаридов в фо-тосинтезирующих листьях из-за снижения интенсивности фотосинтеза, затем количество моносахаридов может возрастать как следствие гидролиза полисахаридов листьев нижних ярусов. При длительном водном дефиците наблюдается уменьшение количества всех форм Сахаров.
Длительный водный дефицит снижает интенсивность фотосинтеза и, как следствие, уменьшает образование АТР в процессе фотосинтетического фосфорилирования. Под влиянием почвенной и атмосферной засухи тормозится также отток продуктов фотосинтеза из листьев в другие органы.
В условиях оптимального водоснабжения наблюдается положительная корреляция между интенсивностью дыхания и количеством фосфорилированных продуктов. Водный дефицит по-разному сказывается на дыхании листьев разного возраста: в молодых листьях содержание фосфорилированных продуктов резко падает, как и интенсивность дыхания, а у листьев, закончивших рост, эта разница четко не проявляется. При дефиците воды снижается дыхательный коэффициент.
 Необходимо отметить, что в условиях водного дефицита верхние листья, в которых за счет некоторого усиления гидролитических процессов увеличивается содержание осмотически активных веществ, оттягивают воду от нижних листьев и дольше сохраняют ненарушенными синтетические процессы, а нижние листья в этих условиях засыхают раньше верхних.
Необходимо отметить, что в условиях водного дефицита верхние листья, в которых за счет некоторого усиления гидролитических процессов увеличивается содержание осмотически активных веществ, оттягивают воду от нижних листьев и дольше сохраняют ненарушенными синтетические процессы, а нижние листья в этих условиях засыхают раньше верхних.
Очевидно, в аридных и близких к ним зонах для сельскохозяйственных культур важно знать физиологические показатели, характеризующие водный режим тканей, и, пользуясь ими, определять срок полива и его продолжительность.
Общеизвестно, что периодически повторяющиеся в некоторых районах СССР засухи и суховеи наносят огромный ущерб сельскому хозяйству. В настоящее время разработана система мероприятий по борьбе с засухой, в числе которых большое место занимает искусственное орошение. Суть его заключается в установлении рационального поливного режима и системы питания растений в разных почвенно-климатических районах.
Среди многочисленных проблем поливного режима сельскохозяйственных культур наиболее важен вопрос о верхней и нижней границах допустимой влажности почвы в активном корнеобитаемом слое.
Как уже отмечалось, верхний предел доступной почвенной влаги называют полевой влагоемкостью. Если влажность почвы превышает полевую влагоемкость, то избыточная влага, несмотря на ее доступность, оказывается малополезной для растений из-за недостатка кислорода. При иссушении почвы до влажности устойчивого завядания (нижний предел доступной влаги) растение страдает от острой почвенной засухи, неблагоприятно сказывающейся на всей его дальнейшей жизнедеятельности, в том числе и величине урожая.
Влажность устойчивого завядания не может служить расчетным и диагностирующим показателем для определения сроков и норм полива, так как снижение урожайности при почвенной засухе вызывается в основном не завяданием растений, а постоянным водным дефицитом в их тканях. Этот дефицит появляется в растениях при такой влажности почвы, при которой ее водоудерживающая сила препятствует полному выравниванию водного дефицита.
Несмотря на то что вода в интервале от полевой влагоем-кости до влажности устойчивого завядания для растений практически доступна, степень ее доступности в пределах указанного интервала неодинакова. В практике орошаемого земледелия очень важно правильно определить то состояние влажности почвы, ниже которой замедляются и нарушаются нормальные физиологические процессы, уменьшается накопление органической массы и снижается урожай. Такое состояние влажности почвы называют нижним пределом оптимальной влажности.
Потребность растений в воде в течение вегетации, а следовательно, величина нижнего предела оптимальной влажности почвы зависит от фаз развития растения, особенностей почвен-но-гидрологических (плодородие почвы, засоление и минерализация грунтовых вод) и климатических условий, уровня агротехники и запланированной величины урожая, а также от наличия или отсутствия критических периодов, характеризующихся острым недостатком воды. Так, для хлопчатника на типичных сероземах эти величины составляют: до цветения—75% полевой влагоемкости, в период цветения — плодо-образования — 70, в период созревания урожая — 60% от полевой влагоемкости. В зависимости от особенностей сорта этот уровень может несколько отклоняться в сторону более высокой влажности для скороспелых сортов и в сторону пониженной влажности для позднеспелых сортов.
Однако в производственных условиях определение сроков полива по влажности почвы очень трудоемко, хотя и применяется для горизонта почвы на глубине 30—40 см. Разработаны методы определения сроков полива исходя из состояния растения: уровня оводненности листьев, их осмотического давления, степени отверстости устьиц и способности выделять пасоку. Наибольшее распространение получил метод определения сосущей силы листьев в разных условиях водного режима растений. В настоящее время известны для различных культур критические величины сосущей силы листьев, коррелирующие с тем состоянием водного режима растений, которое еще не вызывает нарушений обмена веществ, и проведенный вовремя полив обычно приводит к повышению урожая. Например, величины сосущей силы листьев яровой пшеницы в фазу кущения — трубкования равны 0,81—0,91 МПа, в период трубкования— колошения — 1,0— 1,1 и 1,11 — 1,21 МПа — в период налива зерна. Разработка физиологических показателей, позволяющих наиболее точно и экономно поливать растения в условиях засушливых сельскохозяйственных зон,—обязательный элемент высокой культуры орошаемого земледелия.
Орошение — один из приемов мелиорации (радикального улучшения почв). Для заболоченных почв мелиорация состоит в осушении и известковании, для засоленных — в гипсовании и промывке (см. 14.5).
Вода составляет 70 — 95% массы растения. Поступление ее в клетку определяется величиной сосущей силы (водного потенциала — Ψн2о), зависящей от осмотического давления (Ψπ) вакуолярного сока и тургорного (гидростатического) давления (ΨP) клеточного содержимого.
Осмотически поглощенная клетками корня вода движется ради-ально через корень как по апопласту, так и по симпласту до сосудов ксилемы. По ксилеме транспорт воды осуществляется с помощью корневого давления (нижний концевой двигатель) и тран-спирации (верхний концевой двигатель). Движущей силой восходящего тока воды по ксилеме является градиент водного потенциала через растение от почвы до атмосферы. Большая часть поглощенной воды испаряется в процессе транспирации, а около 0,2 % используется на поддержание тургора клеток, метаболизм, транспорт органических веществ по флоэме. Дефицит влаги в клетках влияет на все физиологические процессы, снижая продуктивность растений.
Глава 6

«...По мере расширения работ, связанных с применением удобрений, потребность к установлении принципиальных отправных положений по вопросам минерального питания становится все более настоятельной».
Д. А. Сабинин

Представления о почвенном питании растений начали складываться в связи с развитием растениеводства. Уже в VI —V тысячелетии до новой эры возделывались пшеница, ячмень, рожь, кукуруза, лен, конопля, многие огородные культуры и плодовые деревья, а зола, ил и навоз использовались как средства, повышающие плодородие почвы.
 Первый физиологический эксперимент с целью изучения питания растений был проведен голландским естествоиспытателем Я. Б. ван Гельмонтом в 1629 г. Он посадил в глиняный сосуд, содержащий 91 кг сухой почвы, ивовую ветвь массой 2,25 кг и регулярно поливал почву дождевой водой. Через 5 лет растение и почва были взвешены отдельно. Оказалось, что ива весила 77 кг (прибавка около 75 кг), а масса сухой почвы уменьшилась всего на 56,6 г. Таким образом, масса растения увеличилась в 33 раза, не считая ежегодно опадавших листьев. Ван Гельмонт сделал вывод, что вся растительная масса была создана за счет воды, вносившейся в сосуд при поливе. Этот опыт послужил основой для «водной теории» питания растений, которая довольно долго держалась в ботанике.
Первый физиологический эксперимент с целью изучения питания растений был проведен голландским естествоиспытателем Я. Б. ван Гельмонтом в 1629 г. Он посадил в глиняный сосуд, содержащий 91 кг сухой почвы, ивовую ветвь массой 2,25 кг и регулярно поливал почву дождевой водой. Через 5 лет растение и почва были взвешены отдельно. Оказалось, что ива весила 77 кг (прибавка около 75 кг), а масса сухой почвы уменьшилась всего на 56,6 г. Таким образом, масса растения увеличилась в 33 раза, не считая ежегодно опадавших листьев. Ван Гельмонт сделал вывод, что вся растительная масса была создана за счет воды, вносившейся в сосуд при поливе. Этот опыт послужил основой для «водной теории» питания растений, которая довольно долго держалась в ботанике.
Значительно раньше Аристотелем (384—322 гг. до н. э.) было выдвинуто представление о том, что растения поглощают пищу из почвы в виде сложных веществ. Это представление в конце XVIII - начале XIX в. было развито немецким агрономом А. Тэером, который разработал «гумусовую теорию» питания растений. Давно уже было известно, что чем темнее почва, тем она плодороднее. Темный цвет почвы зависит от содержания в ней различных органических остатков, или гумуса (перегноя). Согласно гумусовой теории, растения питаются водой и гумусом.
Однако постепенно накапливались данные о роли минеральных элементов в питании растений. Один из основоположников отечественной агрономии А. Т. Болотов наметил основные принципы минерального питания растений. В 1770 г. вышла его книга «Об удобрении земель» — первая русская монография по агрохимии. В ней он писал, что пища растений в почве «состоит в воде и некоторых особлевых земляных или паче минеральных частичках...». Болотов разработал приемы внесения удобрений в почву и в одной из статей перечислил 53 вида удобрений, пригодных для применения в сельском хозяйстве.
Швейцарский естествоиспытатель Н. Т. Соссюр систематизировал известные в то время данные о питании растений и установил, что почва снабжает растения азотом и минеральными элементами. При выращивании растений из семян только на дистиллированной воде прирост золы не наблюдался. В своем труде «Химические исследования растений» (1804) он обратил внимание на то, что различные соли поглощаются корнями из водного раствора с неодинаковой скоростью. Французский агрохимик Ж. Б. Буссенго (1837) показал, что растения можно выращивать к на чистом песке, если вносить в него минеральные соли (золу и селитру).
 Немецкий химик Ю. Либих — один из основателей агрохимии, возражая против гумусовой теории, в 1840 г. опубликовал книгу «Химия в приложении к земледелию и физиологии», где обосновал теорию минерального питания растений. По этой теории основой плодородия являются минеральные вещества почвы. Либих считал, что перегной нужен лишь для образования СО2, который ускоряет выветривание материнской горной породы и увеличивает культурный слой почвы. Он первым предложил вносить в качестве удобрений чистые минеральные вещества. Правильно оценивая значение минеральных элементов в питании растений, Либих в то же время считал, что растения поглощают азот из воздуха в виде аммиака. Лишь позднее, в 1856 г., под давлением неопровержимых фактов Либих вынужден был признать, что источником азота для минерального питания растений могут быть нитраты. Неверным в теории Либиха было также отрицание роли органических веществ почвы для развития растения. Как теперь известно, гумус необходим не столько для развития самого растения, сколько для микрофлоры почвы, которая играет весьма значительную роль в питании растений.
Немецкий химик Ю. Либих — один из основателей агрохимии, возражая против гумусовой теории, в 1840 г. опубликовал книгу «Химия в приложении к земледелию и физиологии», где обосновал теорию минерального питания растений. По этой теории основой плодородия являются минеральные вещества почвы. Либих считал, что перегной нужен лишь для образования СО2, который ускоряет выветривание материнской горной породы и увеличивает культурный слой почвы. Он первым предложил вносить в качестве удобрений чистые минеральные вещества. Правильно оценивая значение минеральных элементов в питании растений, Либих в то же время считал, что растения поглощают азот из воздуха в виде аммиака. Лишь позднее, в 1856 г., под давлением неопровержимых фактов Либих вынужден был признать, что источником азота для минерального питания растений могут быть нитраты. Неверным в теории Либиха было также отрицание роли органических веществ почвы для развития растения. Как теперь известно, гумус необходим не столько для развития самого растения, сколько для микрофлоры почвы, которая играет весьма значительную роль в питании растений.
Либих сформулировал «закон минимума», согласно которому внесение любого количества минеральных веществ не даст прироста урожая, пока не будет ликвидирован недостаток веществ, содержащихся в минимальном количестве, а также «закон возврата», указывающий на необходимость возврата в почву питательных веществ, поглощенных растениями. Он утверждал, что несоблюдение принципа полного возврата должно привести к истощению почвы и падению ее плодородия. В целом эти положения верны, но некоторые последователи Либиха вывели из них «закон убывающего плодородия почв». Практика современного земледелия показала полную несостоятельность этого псевдозакона. Урожайность сельскохозяйственных культур при правильной агротехнике и внесении минеральных удобрений постоянно возрастает.
Окончательно опровергли «гумусовую теорию» опыты И. Кнопа и Ю. Сакса (1859). Они показали, что вполне возможно вырастить нормальное растение на воде до полного созревания при его обеспечении лишь семью элементами: азо-
 том, фосфором, серой, калием, кальцием, магнием и железом. Эти опыты окончательно утвердили теорию минерального питания и создали основу для использования вегетационного метода, в том числе водных и песчаных культур. Питательный раствор, разработанный Кнопом, применяется до сих пор.
том, фосфором, серой, калием, кальцием, магнием и железом. Эти опыты окончательно утвердили теорию минерального питания и создали основу для использования вегетационного метода, в том числе водных и песчаных культур. Питательный раствор, разработанный Кнопом, применяется до сих пор.
Буссенго, используя метод вегетационных сосудов и точные количественные измерения, показал, что высшие растения, как правило, не могут связывать атмосферный азот. Этой способностью обладают бобовые растения. Таким образом было впервые установлено, что бобовые растения в отличие от большинства других способствуют накоплению азота в почве. Немецкий ботаник и микробиолог Г. Гельригель в 1880 г. показал, что бобовые растения осуществляют азотфиксацию в симбиозе с клубеньковыми бактериями. Сами бактерии в клубеньках бобовых впервые были обнаружены русским ботаником М. С. Ворониным в 1866 г.
Обширные исследования биологических процессов, происходящих в почве, провел С. Н. Виноградский, который по праву считается основателем почвенной микробиологии. В настоящее время известно, что в почве обитают самые разнообразные микроорганизмы: 1) аммонификаторы, разлагающие органические азотистые соединения (белки, нуклеиновые кислоты, мочевину и др.) с выделением аммиака; 2) азотфиксаторы — микроорганизмы, связывающие молекулярный азот; 3) нитрифи-каторы, которые, используя кислород, окисляют аммиак до нитратов; 4) денитрификаторы, превращающие нитраты в молекулярный азот. При недостатке О2 денитрификаторы используют кислород нитратов и тем самым обедняют почву, возвращая азот в атмосферу.
Кроме бактерий, деятельность которых связана с трансформацией форм азота, в почве имеются бактерии, разлагающие целлюлозу, перерабатывающие различные соединения серы и фосфора, силикатные бактерии, участвующие в освобождении калия из силикатов почв, и др. Некоторые микроорганизмы снабжают растения витаминами, аминокислотами, поэтому роль микроорганизмов почв значительно шире и сложнее, чем просто участие в обмене минеральных веществ.
 Русские ученые П. А. Костычев и В. В. Докучаев разработали основы научного почвоведения. Советский агрохимик К. К. Гедройц обосновал учение о почвенном поглощающем комплексе. Вещества, в том числе и минеральные, удерживаются в почве различными способами: механическим путем, физическими взаимодействиями, химическим и биологическим связыванием веществ. Особое значение Гедройц придавал физико-химической, или обменной, адсорбции, которая осуществляется неорганической (цеолитной) и органической (гумусовой) компонентами почвы. Он установил, что в обменной адсорбции большую роль играют кислые группы как органической, так и неорганической (силикатные группы) части почвы. В основном почвы обладают свойствами катионообменников, хотя в них есть и анионсвязывающие группы.
Русские ученые П. А. Костычев и В. В. Докучаев разработали основы научного почвоведения. Советский агрохимик К. К. Гедройц обосновал учение о почвенном поглощающем комплексе. Вещества, в том числе и минеральные, удерживаются в почве различными способами: механическим путем, физическими взаимодействиями, химическим и биологическим связыванием веществ. Особое значение Гедройц придавал физико-химической, или обменной, адсорбции, которая осуществляется неорганической (цеолитной) и органической (гумусовой) компонентами почвы. Он установил, что в обменной адсорбции большую роль играют кислые группы как органической, так и неорганической (силикатные группы) части почвы. В основном почвы обладают свойствами катионообменников, хотя в них есть и анионсвязывающие группы.
Все эти исследования привели к ясному пониманию того, что плодородие почв связано как со специфическими особенностями материнской горной породы (минеральный состав и структурное состояние почвы), так и с деятельностью почвенных микроорганизмов, которые минерализуют органические остатки.
Растения способны поглощать из окружающей среды в больших или меньших количествах практически все элементы периодической системы. Между тем для нормального жизненного цикла растительного организма необходима лишь определенная группа основных питательных элементов, функции которых в растении не могут быть заменены другими химическими элементами. В эту группу входят следующие 19 элементов:
 Углерод С Калий К Цинк Zn
Углерод С Калий К Цинк Zn
Водород Н Кальций Са Молибден Мо
Кислород О Магний Mg Бор В
Азот N Железо Fe Хлор С1
Фосфор Р Марганец Мп (Натрий) Na
Сера S Медь Си (Кремний) Si
(Кобальт) Со
Среди этих основных питательных элементов лишь 16 являются собственно минеральными, так как С, Н и О поступают в растения преимущественно в виде СО2, О2 и Н2О. Элементы Na, Si и Со приведены в скобках, поскольку их необходимость для всех высших растений пока не установлена. Натрий поглощается в относительно высоких количествах некоторыми видамисем. Chenopodiaceae (маревых), в частности свеклой, а также видами, адаптированными к условиям засоления, и в этом случае является необходимым. То же справедливо для кремния, который в особенно больших количествах встречается в соломине злаковых, для риса он является необходимым элементом.
Первые четыре элемента — С, Н, О, N — называют органогенами. Углерод в среднем составляет 45% сухой массы тканей, кислород — 42, водород — 6,5 и азот— 1,5, а все вместе—95%. Оставшиеся 5% приходятся на зольные вещества: Р, S, К, Са, Mg, Fe, Al, Si, Na и др. О минеральном составе растений обычно судят по анализу золы, остающейся после сжигания органического вещества растений. Содержание минеральных элементов (или их окислов) в растении выражают, как правило, в процентах по отношению к массе сухого вещества или в процентах к массе золы. Перечисленные выше вещества золы относят к макроэлементам.
Элементы, которые присутствуют в тканях в концентрациях 0,001 % и ниже от сухой массы тканей, называют микроэлементами. Некоторые из них играют важную роль в обмене веществ (Мп, Сu, Zn, Со, Мо, В, С1).
Содержание того или другого элемента в тканях растений непостоянно и может сильно изменяться под влиянием факторов внешней среды. Например, Al, Ni, F и другие могут накапливаться в растениях до токсического уровня. Среди высших растений встречаются виды, резко различающиеся по содержанию в тканях таких элементов, как Na, о чем уже говорилось, и Са, в связи с чем выделяют группы растений натриефилов, калъциефилов (большинство бобовых, в том числе фасоль, бобы, клевер), калъциефобов (люпин, белоус, щавелек и др.). Эти видовые особенности обусловлены характером почв в местах происхождения и обитания видов, определенной генетически закрепленной ролью, которую указанные элементы играют в обмене веществ растений.
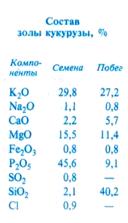
 Наиболее богаты минеральными элементами листья, у которых зола может составлять от 2 до 15% от массы сухого вещества. Минимальное содержание золы (0,4—1%) обнаружено в стволах древесных.
Наиболее богаты минеральными элементами листья, у которых зола может составлять от 2 до 15% от массы сухого вещества. Минимальное содержание золы (0,4—1%) обнаружено в стволах древесных.
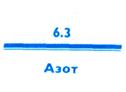
Азот был открыт в 1772 г. шотландским химиком, ботаником и врачом Д. Резерфордом как газ, не поддерживающий дыхание и горение. Поэтому он и был назван азотом, что значит «нежизненный». Однако азот входит в состав белков, нуклеиновых кислот и многих жизненно важных органических веществ. Ликвидация недостатка некоторых незаменимых азотсодержащих соединений — аминокислот, витаминов и др. — наиболее острая проблема продовольственных программ человечества.
 Для растений азот — дефицитный элемент. Если некоторые микроорганизмы способны усваивать атмосферный азот, то растения могут использовать лишь азот минеральный, а животные — только азот органического происхождения, да и то не любой. Например, мочевина животным организмом непосредственно не усваивается. В то время как животные относятся к азоту довольно расточительно, выделяя мочевую кислоту, мочевину и другие азотсодержащие вещества, растения почти никогда не выделяют азотистые соединения как продукты отброса и там, где это возможно, азотистые соединения заменены на безазотистые вещества. Например, у растений в состав полисахаридов клеточных оболочек не входят гексоза-мины, характерные для мукополисахаридов животных и хитина членистоногих и грибов.
Для растений азот — дефицитный элемент. Если некоторые микроорганизмы способны усваивать атмосферный азот, то растения могут использовать лишь азот минеральный, а животные — только азот органического происхождения, да и то не любой. Например, мочевина животным организмом непосредственно не усваивается. В то время как животные относятся к азоту довольно расточительно, выделяя мочевую кислоту, мочевину и другие азотсодержащие вещества, растения почти никогда не выделяют азотистые соединения как продукты отброса и там, где это возможно, азотистые соединения заменены на безазотистые вещества. Например, у растений в состав полисахаридов клеточных оболочек не входят гексоза-мины, характерные для мукополисахаридов животных и хитина членистоногих и грибов.

