 2015-06-10
2015-06-10 1025
1025Промежуточный мозг (diencephalon) располагается в центре головного мозга между двумя полушариями. Его можно разделить на отделы,
 |
 |
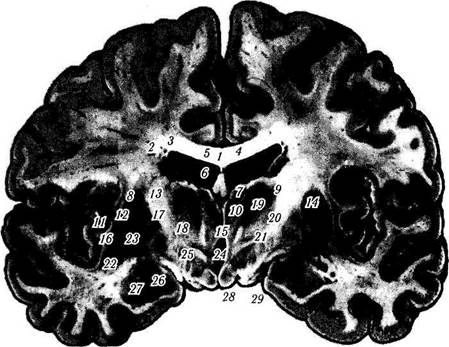
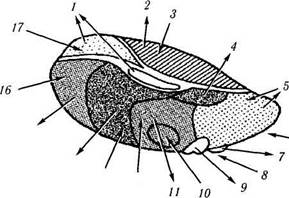
Рис. 4.1.17. Локализация ядер зрительного бугра:
а — ядерный парамагнитный резонанс; б — фронтальный срез головного мозга (/ — тело мозолистого тела; 2 — тело хвостатого ядра; 3 — центральная часть бокового желудочка; 4 — прозрачная перегородка; 5 — сосудистое сплетение бокового желудочка; 6 — свод; 7 — передняя группа ядер зрительного бугра; 8 — наружная капсула; 9 — латеральная группа ядер зрительного бугра; 10 — медиальные ядра зрительного бугра; //— скорлупа; 12 — латеральная медуллярная пластинка; 13 — внутренняя капсула; 14 — ретикулярное ядро зрительного бугра; 15 — межталамическая спайка; 16 — латеральная часть бледного шара; 17 — медиальная медуллярная пластинка; 18 — мамиллоталамический пучок; 19 — HI поле Фореля; 20 — гопа incerta; 21 — Н2 поле Фореля; 22 — ограда; 23 —медиальная часть бледного шара; 24 — третий желудочек; 25 — ядра гипоталамуса; 26 —зрительный тракт; 27 — миндалевидное тело; 28 —сосковидное тело; 29 —основание ножек мозга)
Анатомия головного мозга

 которые анатомически в той или иной степени взаимосвязаны [4, 6, 8, 9]:
которые анатомически в той или иной степени взаимосвязаны [4, 6, 8, 9]:
1. Зрительный бугор (thalamus).
2. Забугорная область (metathalamus).
3. Подбугорная область (hypothalamus).
4. Надбугорная область (epithalamus).
5. Субталамическая область (subthalamus).
Зрительный бугор (thalamus) (рис. 4.1.17—
4.1.20).
Функциональное значение зрительного бугра велико. Таламус можно считать воротами для поступления сенсорной информации от всех систем организма к филогенетически более молодым церебральным структурам, которые делают возможным целенаправленное, сознательное поведение [565].
Зрительный бугор состоит из скоплений ядер, отличающихся своими функциональными особенностями. Ряд авторов в соответствии с функцией ядер зрительного бугра разделяет их на четыре группы:
1. Специфические ядра переключения для
соматосенсорной, зрительной и слуховой аффе
рентных систем.
2. Ядра неспецифической системы.
3. Ядра с преимущественно двигательными
функциями.
4. Ядра с ассоциативными функциями. Боль
шая часть этих ядер связана с корой мозга
мощными путями.
Анатомически зрительный бугор представляет собой два больших овоидной формы скопления ганглиозных клеток, расположенных выше ножек мозга по бокам третьего желудочка [4, 6—9, 11, 398, 578]. Передне-задний размер каждого образования равен примерно 4 см, а ширина и высота — 2,5 см (рис. 4.1.19, 4.1.20).
Передняя часть зрительного бугра узкая и располагается вблизи средней линии. Задней границей, является межжелудочковое отверстие. Расширенный задний полюс или подушка накладывается на верхнее четверохолмие.
Снизу и медиальней подушки зрительного бугра лежит медиальное коленчатое тело. С латеральной стороны зрительный бугор отделен от чечевицеобразного ядра (lenticular nucleus) полосатым телом (corpus striatum) и задней частью внутренней капсулы (capsula interna).
В специфических ядрах органов чувств происходит переключение определенной сенсорной информации. К этим ядрам относятся медиальное и латеральное коленчатые тела. Медиальное коленчатое тело участвует в обработке слуховой информации и проецируется на специфические участки коры мозга (слуховое поле). Латеральное коленчатое тело определяет функцию зрительного восприятия, относится к зрительному пути. По этой причине оно подробно будет описано в следующих разделах.
Еще одно ядро, которое можно рассматривать как проекционный центр для специфичес-
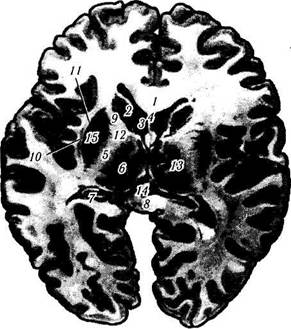
Рис. 4.1.18. Горизонтальный срез головного мозга через ядра зрительного бугра:
 |
| 12 |
/ — колено мозолистого тела; 2 — головка хвостатого ядра; 3 — фронтальный рог бокового желудочка; 4 — прозрачная перегородка; 5 — внутренняя капсула; 6 — зрительный бугор; 7 — ножка свода; 8 — утолщение (валик) мозолистого тела; 9 — передний край внутренней капсулы; 10 — ограда; // — наружная капсула; 12 — колено внутренней капсулы; 13 — скорлупа; 14 — шишковидная железа; /5 — чечевицеобразное ядро
Рис. 4.1.19. Схематическое изображение расположения
ядер, путей и пучков зрительного бугра (по Heinz
Feneis, 1994):
1 — заднее латеральное ядро; 2 — дорзальное латеральное ядро; 3 — переднее вентральное ядро; 4 — вентральное промежуточное ядро; 5 — медиальное вентральное ядро; 6 — заднелатераль-ное вентральное ядро; 7 — заднемедиальное вентральное ядро; 8 — ядра подушки; 9 — латеральное (наружное) коленчатое тело; 10 — медиальное коленчатое тело; //—латеральная петля (lem-niscus lateralis); 12 — медиальная петля (lemniscus medialis); 13 — спинномозговая петля (lemniscus spinalis); 14 — тройничная петля (lemniscus trigeminalis); 15 — слуховая лучистость; 16 — зрительная (Грасиоле) лучистость; 17 — медиальные ядра; 18 — передние ядра
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

 |
| 15 |
| 14 |
| 13 |
| 12 |
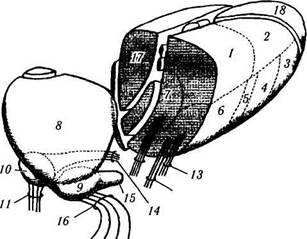
Рис. 4.1.20. Основные афферентные и эфферентные проекции зрительного бугра (по Carpenter, 1976):
I — поясная извилина; 2 — прелобная кора; 3 — миндалевидный комплекс, височная кора; 4 — верхняя теменная доля; 5 — поля 18 и 19, нижняя теменная доля; 6 — нижние бугорки; 7— латеральный лемниск; 8 — зрительный тракт; 9 — поле 17; 10 — тригеминоталамический тракт; // — поля 3, 1 и 2; 12 — медиальный лемниск, спинноталамический тракт; 13 — поле 4; 14 — зубчатое ядро, бледный шар, черная субстанция; /5 — поле 6, лобная кора; 16 — бледный шар, черная субстанция; 17 — ма-миллоталамический тракт, свод
кой области коры, — это переднее ядро зрительного бугра (п. anterior thalami). Через это ядро проходит информация от сосцевидного тела (corpus mammilare) к передним отделам поясной извилины (рис. 4.1.17—4.1.20).
Специфическим таламическим ядром сомато-сенсорной системы является вентробазальное ядро или вентробазальный комплекс. Оно делится на вентральное заднелатеральное ядро (п. ventralis posteriolateralis) и вентральное заднемедиальное ядро (п. ventralis posterio-medialis). Нейронное представительство туловища лежит в вентральном заднелатеральном ядре, а лица — в вентральном заднемедиаль-ном. К вентральному заднелатеральному ядру ведут медиальный лемниск и неоспинноталами-ческий тракт, а к вентральному заднемедиаль-ному ядру приходят соответствующие пути от ядра тройничного нерва. Вентробазальное ядро является вторым переключением в лемниско-вой системе.
Вторая группа ядер формирует менее четко организованные структурные образования. Относятся они к неспецифическим проекционным ядрам. Главными среди неспецифических ядер являются медиально-расположенные участки, граничащие с третьим желудочком, и интрала-минарные ядра. Они функционально связаны с ретикулярной формацией ствола мозга и, подобно ей, получают афферентацию от всех органов чувств [240]. Считается также, что неспецифические таламические ядра осуществляют восприятие и оценку болевых стимулов. Неспецифические ядра таламуса являются вторичными структурами переключения, которые распределяют афферентную информацию, проецирующуюся на ретикулярную формацию.
Как указано выше, в зрительном бугре существуют важные двигательные ядра. К подобным ядрам относятся п. ventralis posterior — medialis, et lateralis, промежуточное вентральное ядро (п. ventralis thalami inter-medialis). В самых ростральных отделах вентральной группы ядер находится переднее вентральное ядро зрительного бугра (п. ventralis thalami anterior). Именно эти ядра соединяют мозжечок и базальные ганглии с двигательной корой. Импульсы, исходящие из этих ядер, оказывают модифицирующее влияние на двигательные стимулы, посылаемые в направлении мышц.
Другие ядра зрительного бугра выполняют ассоциативную функцию. Термин «ассоциативное ядро» применяется к тем таламическим ядрам, которые обладают отчетливыми связями с областями коры, но не могут быть отнесены к какой-либо сенсорной системе. Они принимают участие в высших интегративных процессах в головном мозге. К представителям этой группы относятся три ядра, каждое из которых связано с одной из главных ассоциативных областей коры. Это подушка зрительного бугра (связана с ассоциативной зоной теменной и височной коры), заднее латеральное ядро (п. lateralis posterior) (связано с теменной корой) и медиальное дорзальное ядро (п. medialis dorsalis) (связано с лобной долей).
Латеральные ядра зрительного бугра, к которым относится и подушка зрительного бугра, отвечают за болевую чувствительность. Она также имеет обширные связи с затылочной долей головного мозга, включая зрительную кору. В пределах подушки лежат нейроны, отвечающие за анализ различных характеристик изображения, например цвета, определения направления цели и ее ориентацию [23, 43, 50, 192, 287, 341, 365, 407, 448, 561].
В настоящее время предполагают, что подушка обеспечивает [96, 201, 448, 449, 558, 561]:
1) выбор цели «зрительного интереса»;
2) «фильтрацию» зрительных стимулов;
3) использование информации «зрительного
интереса» при определении направления дви
жения глаза или других частей тела.
В переднем отделе подушки выявлены ре-тинотопические поля [44], содержащие полную карту сетчатой оболочки противоположного глаза («вторая приводящая зрительная система» — коликуло-пульвинар-париетальный тракт). Ретинотопические области отвечают за саккадические движения глаз. На эту область подушки проецируются верхнее четверохолмие, зрительная кора и другие области коры головного мозга (префронтальная кора, задняя часть теменной и верхней височной коры). Существуют и обратные проекции [52, 447].
В дорзомедиальной части латерального отдела подушки выявлена небольшая область, свя-
Анатомия головного мозга

 занная со средней височной областью (область коры 7) и с теменно-затылочной областью.
занная со средней височной областью (область коры 7) и с теменно-затылочной областью.
Нейроны подушки отвечают за пространственное зрительное внимание, о чем свидетельствует нарастание их активности во время появления внимания к зрительному стимулу, расположенному по периферии. Отбор информации, обеспечивающий пространственное внимание, свойственен также клеткам 7-го поля теменной коры головного мозга [416, 595].
Нейроны заднелатеральных полей реагируют только в тех случаях, когда стимулом является зрительная информация, индуцирующая движение глаз [595]. Приблизительно 30% клеток таламуса, реагируют и после прекращения движения глаз. Именно эти клетки предупреждают о необходимости повторного просмотра объекта.
На нейроны заднелатеральных ядер проецируются поверхностные слои верхнего четверохолмия [49, 229, 413, 430, 450]. Благодаря этому зрительномоторные свойства клеток таламуса являются результатом совместной деятельности верхнего четверохолмия и геникуло-стриарного пути.
При повреждении головного мозга в области зрительного бугра возникает нарушение чувствительности различной локализации и степени выраженности, спонтанные боли и др. Важно отметить, что кровоизлияния, локализующиеся в области зрительного бугра, вызывают симптомы, аналогичные симптомам, возникающим при повреждении теменной доли. При этом больные невнимательны к зрительной цели, расположенной в контрлатеральной зрительной области.
Забугорная область (metathalamus). К забугорной области относятся два ядерных образования — латеральное и медиальное коленчатые тела (nuclei geniculati lateralis et medialis) [3, 4, 6, 8, 9]. Латеральное коленчатое тело связано с передачей зрительной информации, а медиальное — лежит на пути проведения слуховой информации.
Подбугорная область, или гипоталамус (hypothalamus) (рис. 4.1.21, 4.1.22). Гипоталамус является наиболее ростральным компонентом автономной (вегетативной, или висцеральной) нервной системы. Содержит он особые нейросекреторные ядра, клетки которых вырабатывают и секретируют в кровь нейрогормоны [3, 9, 11, 397, 417].
Эти клетки получают афферентные импульсы из других частей нервной системы, а их аксоны оканчиваются на кровеносных сосудах (аксо-вазальные синапсы).
Гипоталамус формирует передние и боковые стенки третьего желудочка. Его верхняя граница отделена от зрительного бугра гипоталами-ческой бороздой. Нижней границей гипоталамуса служит средний мозг, а верхней — конечная пластинка, передняя спайка и зрительный пере-
крест. Латеральнее гипоталамуса расположены зрительный тракт, внутренняя капсула и субта-ламические структуры.
В гипоталамусе выделяют три группы ядер: передняя, средняя и задняя.
К передней группе ядер относят надзри-тельное (супраоптическое; п. supraopticus) и паравентрикулярное (п. paraventricularis) ядра. Надзрительное ядро находится сбоку и выше зрительного перекреста и состоит приблизительно из 750 000 нейронов, относящихся к парасимпатической системе. Это ядро участвует в циркадных циклах [481].
Паравентрикулярное ядро лежит сразу же под эпендимой передней стенки третьего желудочка и состоит из 55 000 нейронов.
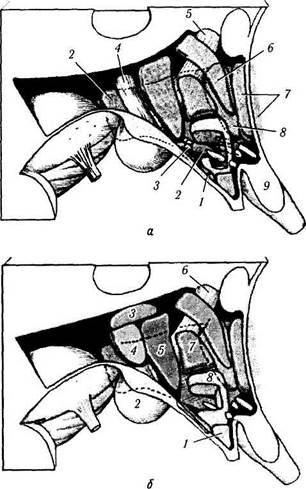
Рис. 4.1.21. Схематическое изображение расположения ядер гипоталамуса (по Heinz Feneis, 1994):
а: I —дугообразное ядро; 2 — латеральная гипоталамическая область; 3 — серобугорные ядра; 4 — сосцевидно-таламический пучок; 5 — свод; 6 — паравентрикулярные ядра; 7 — предопти-ческое медиальное/латеральное ядро; 8 — переднее гипоталами-ческое ядро; 9 — супраоптическое ядро; б. 1 — ядро воронки (дугообразное ядро); 2 — медиальные и латеральные ядра сосцевидного тела; 3 — дорзальное гипоталамическое ядро; 4 — заднее паравентрикулярное ядро; 5 — заднее гипоталамическое ядро; 6 — свод; 7 — дорзомедиальное гипоталамическое ядро; 8 — вентромедиальное гипоталамическое ядро
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ

 |
| 10 |
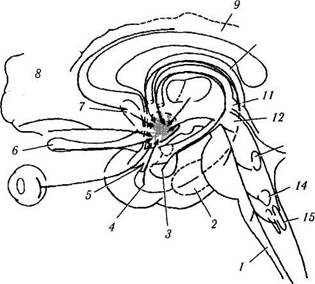
Рис. 4.1.22. Схема афферентных связей подбугорной области (по Brodal. 1992):
1 — ретикулярная формация; 2 — гиппокамп; 3 — миндалина; 4 — обонятельная кора; 5 — зрительный перекрест; 6 — обонятельная луковица; 7 — септальные ядра; 8 — прелобная кора; 9 — поясная извилина; 10 — свод; // — конечная пластинка; 12 — периакведуктальное серое вещество; 13 — nucleus locus coe-ruleus; 14 — ядро шва; 15 — ядро слюноотделительного тракта
К средней группе относят бугорковое, вент-ромедиальное, дорзальное и латеральное ядра.
Задняя группа ядер (regio hypothalamica posterior) располагается вблизи сосцевидных тел и состоит из медиальных и латеральных ядер сосцевидного тела (п. corporis mamillaris medialis, lateralis).
Медиальное ядро складывается из мелких клеток, формирующих толстую связку волокон (сосцевидно-таламический пучок), возникающую в области гиппокампа и непосредственно связанную с передним ядром таламуса с этой же стороны. Группы небольших ядер включают в свой состав надперекрестное ядро, лежащее вблизи срединной линии и выше зрительного перекреста, и дугообразное ядро (п. arcuatus).
Афферентные пути гипоталамуса довольно многочисленны. Некоторые проекции приведены на рис. 4.1.22.
На гипоталамус проецируются также кора большого мозга, бледный шар, миндалина, ядра отдельного тракта, ретикулярная формация, ствол мозга и спинной мозг [4, 6, 8, 9, 74, 240]. У некоторых млекопитающих и человека выявлен ретиногипоталамический путь. Начинается он в небольшой популяции ганглиозных клеток сетчатой оболочки и проецируется на надзрительное ядро [237, 381, 382, 481, 482]. Предполагают, что он обеспечивает фотонейроэндо-кринные и фотопериодические функции, включая циркадные ритмы [311, 481, 563].
К эфферентным путям гипоталамуса относят три четко обозначенных пучка: от сосцевид-
ных тел, от перивентрикулярных ядер и от над-зрительного ядра (п. supraopticus), образующих надзрительно-гипофизарный путь. Направляются они к гипофизу.
Наибольшим путем является сосцевидно-покрышечный путь (/. mammalotegmentalis). Идет он от гипоталамуса к ядрам покрышки среднего мозга и моста. Некоторые волокна также подходят к перегородке, гиппокампу и подушке [4, б, 8, 397].
Волокна могут быть прослежены от вент-ромедиального ядра гипоталамуса к серому веществу среднего мозга, а также к претекталь-ной области, верхним бугоркам, дорсальным и вентральным покрышечным ядрам Гуддена (Gudden), ядрам шва (п. raphes, расположены по средней линии продолговатого мозга) и ядрам locus coeruleus (пигментированное возвышение в верхнем углу основания мозга) [542].
Выявлены эфферентные волокна, направляющиеся к дорзальному двигательному ядру блуждающего нерва, слюноотделительному ядру {п. salivatorius), двойному ядру (п. атЫ-guus, ядро расположено в продолговатом мозге в составе ретикулярной формации и образует двигательные волокна языкоглоточного, блуждающего и добавочного нервов), а также к спинному мозгу [120, 121, 240, 487].
Как было указано выше, гипоталамус посылает большое количество волокон задней доле гипофиза (нейрогипофиз) через надзрительно-гипофизарный путь (tractus supraopticohypo-physialis). У человека этот тракт состоит примерно из 100 000 волокон. Возникают они в над-зрительном и паравентрикулярных ядрах [432].
Необходимо более подробно остановиться на особенностях нейросекреторной функции ядер гипоталамуса. С функциональной точки зрения, ядра гипоталамуса можно разделить на крупно- и мелкоклеточные.
Крупноклеточные ядра образованы клеточными телами, которые в 2—3 раза крупнее, чем в других отделах гипоталамуса. К ним относятся супраоптическое (надзрительное) и паравен-трикулярное ядра. Супраоптическое ядро в 3— 4 раза крупнее паравентрикулярного и лишь в центральных участках состоит из крупных нейронов. Аксоны клеток супраоптического и паравентрикулярного ядер в составе гипоталамо-гипофизарного пути покидают гипоталамус и проникают в заднюю долю гипофиза. При этом волокна пересекают гемато-энцефалический барьер. Образуют они терминали на капиллярах. Крупноклеточные ядра секретируют антидиуретический гормон, или вазопрессин, и окситоцин. Эти гормоны вырабатываются разными клетками. У человека антидиуретический гормон образуется, главным образом, в супраоптическом ядре, а окситоцин — в паравентрикулярном. Эти гормоны синтезируются в гранулярном эндоплазматическом ретикулуме в виде крупных молекул прогормона. Молекулы прогормо-
Анатомия головного мозга

 на переносятся в комплекс Гольджи, где и упаковываются в виде гранул. Процессинг продукта (завершается в ходе транспорта в аксоне) приводит к высвобождению активного гормона и нейрофизина — белка с неясной функцией.
на переносятся в комплекс Гольджи, где и упаковываются в виде гранул. Процессинг продукта (завершается в ходе транспорта в аксоне) приводит к высвобождению активного гормона и нейрофизина — белка с неясной функцией.
Вазопрессин вызывает усиление тонуса гладкомышечных клеток артериол, приводящее к повышению артериального давления. Он также называется антидиуретическим гормоном. Окситоцин вызывает координированные сокращения мышечной оболочки матки во время родов, а также миоэпителиальных клеток в концевых отделах молочной железы.
Мелкоклеточные ядра вырабатывают ряд гипофизарных факторов, которые усиливают (рилизинг-факторы, или либерины) или угнетают (ингибирующие факторы, или статины) выработку гормонов клетками передней доли, попадая к ним по воротной системе сосудов [351].
Аксоны нейросекреторных клеток этих ядер образуют терминали на первичной капиллярной сети в срединном возвышении (нейрогормо-нальной контактной зоне). Эта сеть собирается далее в воротные вены, проникающие в переднюю долю гипофиза, и распадается на вторичную сеть капилляров между рядами железистых клеток — аденоцитов систему [57].
Портальная система гипофиза обеспечивает сосудистую связь между стеблем гипофиза и передней долей, в результате чего передняя доля гипофиза находится под контролем гипоталамуса.
Гипоталамус является координирующим центром вегетативной нервной системы. Он регулирует эмоциональное поведение, половую деятельность, эндокринную секрецию и адаптационные возможности организма при изменении окружающей среды. Кроме того, он осуществляет контроль водного баланса, веса тела, функцию сна и другие соматические реакции.
Задняя часть гипоталамуса обеспечивает деятельность симпатической системы. Стимуляция ее приводит к расширению кровеносных сосудов, повышению температуры, усилению метаболизма и расширению зрачка. Передняя часть гипоталамуса осуществляет контроль над деятельностью парасимпатической системы.
Поражение гипоталамуса приводит к утере способности регулировать температуру тела, снижению половых функций. Следствием поражения гипоталамуса и питуитарной железы является несахарный диабет.
Сосцевидные тела определяют эмоции. Кроме того, они участвуют в процессах памяти. Именно эта область поражена при энцефалопатии Вернике, часто развивающейся у алкоголиков, в результате дефицита тиамина и сопровождается параличом наружных мышц глаза и нистагмом.
Гипофиз (hypophysis, glandula pituitaria). Гипофиз регулирует активность ряда желез внутренней секреции. Как было указано выше,
нейрогипофиз является местом хранения и высвобождения в кровь многих гормонов и биологически активных веществ.
Состоит он из двух эмбриологически, структурно и функционально отличающихся частей — нейрогипофиза (вырост промежуточного мозга) и аденогипофиза [4, б, 8, 9, 11]. Адено-гипофиз разделяется на более крупную переднюю долю, узкую промежуточную и слабо развитую трабекулярную часть. Не описывая подробно строение гипофиза, мы лишь укажем, что хромофильные аденоциты передней доли гипофиза подразделяются на ацидофилы и базофилы. Ацидофилы вырабатывают соматотропный гормон, или гормон роста, пролактин (стимулирует развитие молочных желез и лактацию). Базофилы вырабатывают фолликулостимулиру-ющий, лютеинизирующий, тириотропный, адре-нокортикотропный гормоны.
Промежуточная доля развита слабо и состоит из тяжей базофильных и хромофобных клеток, синтезирующих меланоцитстимулирующий гормон (активирует меланоциты) и липотроп-ный гомон (стимулирует обмен жиров).
Как указывалось выше, задняя доля гипофиза содержит отростки и терминали нейросекреторных клеток супраоптического и паравентри-кулярного ядер. По этим отросткам транспортируются и выделяются в кровь вазопрессин и окситоцин.
Надбугорная область (epithalamus). В состав надбугорнои области входят шишковидное тело, или эпифиз (corpus pineale), и ядра уздечки (habenula), примыкающие с внутренней стороны к дорзальным отделам зрительного бугра [4, 6, 8, 397]. Эта область анатомически связана с лимбической системой и ретикулярной формацией среднего мозга и, по-видимому, осуществляет интеграцию их функций.
У некоторых низших млекопитающих, размножение которых связано со световым режимом года, часть волокон зрительной системы идет в область перегородки или окружающие ее отделы мозга. Импульсы из этих структур достигают затем области уздечки и эпифиза.
Эпифиз (шишковидная железа) является нейроэндокринным органом, получающим информацию из нервной и эндокринной систем. Эта информация интегрируется в нем и регулирует активность пинеалоцитов. У высших позвоночных эпифиз утрачивает фоторецептор-ную функцию и сохраняет гормональную, регулируя циклические процессы в организме. У низших млекопитающих эпифиз функционирует как биологические часы, чувствительные к степени освещения.
Эпифиз состоит из двух типов клеток — светлых и темных пинеалоцитов. Пинеалоциты вырабатывают вещества двух типов: индолами-ны и пептиды. Наиболее важным индоламином является гормон мелатонин, угнетающий секрецию гонадолиберина, снижая активность гонад.
Глава 4. ГОЛОВНОЙ МОЗГ И ГААЗ
 Из нескольких десятков гормонально-активных пептидов, вырабатываемых пинеалоцитами, наиболее важными являются аргинин-вазотоцин, пинеальный антигонадотропный пептид, а также либерины и статины.
Из нескольких десятков гормонально-активных пептидов, вырабатываемых пинеалоцитами, наиболее важными являются аргинин-вазотоцин, пинеальный антигонадотропный пептид, а также либерины и статины.
Для врача офтальмолога шишковидная железа представляет интерес по ряду причин. Во-первых, необходимо помнить о возможности развития опухоли железы — пинеаломы (dys-germinoma). Рост этой опухоли может привести к возникновения синдрома Парино (двухсторонний парез, или паралич взора вверх или вниз, отсутствие или ослабление конвергенции, птоз, нарушение зрачковых реакций, миоз, нередко вертикальный нистагм) из-за близкого расположения претектальной области. Эктопические пинеаломы могут проникать в гипоталамус через третий желудочек. В результате нарушаются различные функции глаза. Сопровождает этот процесс несахарный диабет и преждевременная половая зрелость.
Субталамическая область (subthalamus). К субталамусу относятся ядерное образование с тем же названием, а также несколько менее четко очерченных ядерных областей и пучки волокон, идущие от базальных ядер в конечный мозг. Функционально эта область связана с красным ядром и черным веществом, расположенными в среднем мозге. Все это служит определенным указанием на то, что функция суб-таламической области связана с базальными ядрами.
Экстрапирамидная система. По многим клиническим и физиологическим причинам полезно рассматривать анатомическую область, распространяющуюся от коры головного мозга до мозжечка, как экстрапирамидную систему. Экстрапирамидная система включает базаль-ные ганглии, вентролатеральное ядро таламуса, гипоталамическое ядро промежуточного мозга, ретикулярную формацию, черную субстанцию и красное ядро среднего мозга. В отличие от пирамидной системы экстрапирамидный путь является полисинаптическим. Этот путь взаимодействует с автономной нервной системой, обеспечивая поддержание мышечного тонуса. Выражение лица является наиболее яркой функцией экстрапирамидной системы.
Необходимо рассмотреть также связи, которые экстрапирамидные ядра имеют с другими отделами — головным мозгом, мозжечком и ретикулярной формацией ствола мозга. Самым длинным нисходящим отводящим путем является центральный покрышечно-спинномозговой путь (tractus tecto-spinalis). Идет он от таламуса, базальных ганглиев и красных ядер к нижней маслине спинного мозга. Заканчивается он на нейронах передних рогов спинного мозга. Этот путь контролирует тонус мышц шеи, а также зрительные и слуховые рефлексы. Поражение покрышечно-спинномозгового пути, нижней маслины и зубчатого ядра мозжечка сопро-
вождается глазной и небной миоклонией (гиперкинез, характеризующийся быстрыми кло-ническими подергиваниями мышц или их отдельных пучков, возникающими как в покое, так и при движениях, но исчезающими во сне).
Красноядерно-спинномозговой путь (tractus rubrospinalis) проходит через красное ядро (рис. 4.1.23, см. цв. вкл.). Находится он лате-ральней корково-спинномозгового пути. Начинается он от клеток красного ядра, а заканчивается на нейронах передних рогов спинного мозга. Вдоль этого пути передается информация спинным нервам от мозжечка. Эта информация проходит через верхние мозжечковые ножки, достигает красного ядра и поступает в спинные нервы. Этот тракт имеет большое значение в контроле тонуса поперечнополосатых мышц туловища, поддержании позы.
Ретикулярно-спинномозговой путь (tractus reticulospinalis) начинается в ядрах ретикулярной формации моста и продолговатого мозга. Заканчивается он в передних рогах спинного мозга, участвуя при этом в образовании спинных нервов. Помимо контроля тонуса скелетных мышц, аналогично красноядерно-спинно-мозговому пути, ретикулярно-спинномозговой путь играет значительную роль в контроле автономных функций.
Преддверно-спинномозговой путь (tractus vestibulospinalis) начинается в вестибулярных ядрах, расположенных в мосту и продолговатом мозге, и направляется к мотонейронам передних рогов спинного мозга. Контролирует этот путь равновесие.
Поражение экстрапирамидного пути (бледного шара, черного вещества) приводит к возникновению различных типов дискинезии, или нарушению произвольных движений. Причинами поражения тракта могут явиться дегенеративные, воспалительные (энцефалит) и опухолевые заболевания. Наиболее распространенным дегенеративным заболеванием является болезнь Паркинсона (сочетание гипокинезии и дрожания в покое).
Предполагается, что повреждения черной субстанции вызывает атрофию допаминэрги-ческого нигростриатного тракта. При этом возникает тремор, гипокинетическая дисартрия, ослабление голоса, маскоподобное выражение лица. При повреждении базальных ганглиев развивается хорея. Результатом поражения базальных ганглиев может быть также и спастическая дистония, гиперкинетическая дисартрия.
Через красное ядро проходят волокна от ядер глазодвигательного нерва. По этой причине при разрушении красноядерно-спинномоз-гового пути развивается офтальмоплегия. Повреждение красного ядра может вызвать также контрлатеральную атаксию.
Так как экстрапирамидная система регулирует движения, то не удивительно, что повреждения этой области приводят к нарушению дви-
Анатомия головного мозга
 жения глаз. Необходимо отметить, что в последние годы при экстрапирамидных болезнях используют разрушение вентролатеральных ядер зрительного бугра и бледного шара в процессе стереотаксических операций.
жения глаз. Необходимо отметить, что в последние годы при экстрапирамидных болезнях используют разрушение вентролатеральных ядер зрительного бугра и бледного шара в процессе стереотаксических операций.








