 2015-06-10
2015-06-10 5539
5539Размер зрачка определяется многими факторами. Это возраст, эмоциональное состояние, степень освещения сетчатки, степень аккомодации и др. Изменение диаметра зрачка контролируется действием парасимпатических и симпатических эфферентных трактов.
Зрачковый рефлекс заключается в содружественном и равном сужении зрачков при освещении одного из глаз, обеспечивая при этом уменьшение светового потока, падающего на сетчатку. Сужение зрачка выявляется при исключительно низкой интенсивности освещения и пропорционально интенсивности и продолжительности стимула [344, 548].
Свет, проходя через преломляющие среды глаза, попадает на сетчатку. Фоторецепторы сетчатой оболочки и являются началом рефлекса. Парасимпатическая иннервация сфинктера является эфферентным плечом зрачкового рефлекса рефлекторной дуги.
Афферентный путь (рис. 4.5.10). Афферентный путь начинается в палочках и колбочках сетчатой оболочки и проходит в составе зрительного нерва к структурам центральной нервной системы. До сих пор обсуждается вопрос — «зрительные» и «зрачковые» волокна зрительного нерва идентичны или нет? Если даже волокна «зрачкового рефлекса» являются самостоятельными и не обеспечивают передачу зрительной информации, они все же располагаются вблизи волокон, несущих зрительную информацию. Об этом свидетельствуют факты исчезновения зрачкового рефлекса на слепом глазу (поражение зрительного нерва).
Проходя в зрительном нерве, зрачковые волокна достигают зрительного перекреста, где они частично перекрещиваются и часть их переходит на противоположную сторону.
Затем волокна поступают в зрительный тракт. Повреждение этой области приводит к развитию гемианопсической реакции зрачка Вернике (Wernicke).
В задней трети зрительного тракта, не достигая наружного коленчатого тела, волокна покидают зрительный тракт и проходят поверхностно в составе ручки верхнего бугорка по направлению к латеральной части верхнего холмика четверохолмия [71, 548] (рис. 4.5.10). Разрушение обеих ручек верхнего холмика приводит к тому, что зрачок не реагирует при освещении обоих глаз [290, 291].
Ни одно из волокон зрачкового рефлекторного пути, по-видимому, не заканчивается в наружном коленчатом теле. Однако некоторые исследователи полагают, что возможно переключение части волокон, идущих в претекталь-ную область, в прегеникулярном ядре, хотя морфологическими методами существование подобных связей не установлено.
В последующем «зрачковые» волокна проходят к среднему мозгу по боковой поверхности верхнего четверохолмия и достигают парного претектального ядра (плохо очерченное скопление мелких клеток, расположенных впереди латерального края верхнего четверохолмия). Здесь волокна прерываются, образуя терминалы (рис. 4.5.10, б).
Многочисленные подгруппы нейронов относят к претектальным ядрам, хотя их функциональное значение не совсем ясно. К ним относят ядро оливы, подчечевицеобразное ядро, ядро зрительного тракта, заднее ядро и пред-крышечное ядро (рис. 4.5.11).
Волокна, идущие от сетчатки, заканчиваются преимущественно в дорзомедиальной части ядра оливы (п. olivaris) с этой же стороны, а также в подчечевицеобразном ядре противоположной стороны (п. sublentiformis). Аналогичная проекция выявляется и на ядро пред-крышечной области [51, 419, 552].
Аксоны нейронов ядра оливы и подчечевице-подобного ядра частично перекрещиваются
Вегетативная (автономная) иннервация глаза

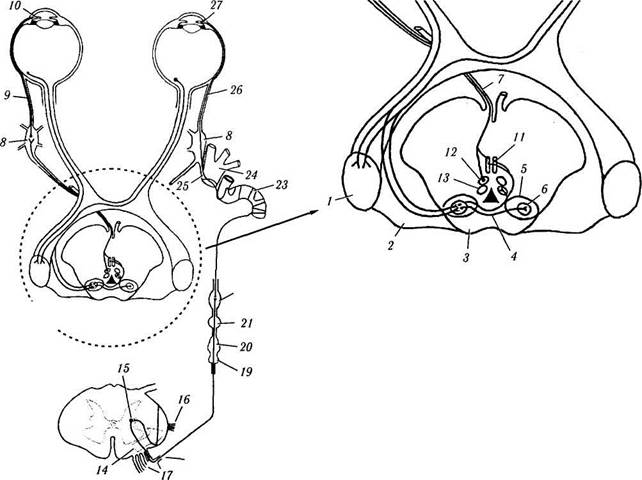 |
| Рис. 4.5.10. Схема иннервации сфинктера и дилятатора радужной оболочки глаза: / — наружное коленчатое тело; 2 — плечо верхних бугорков; 3 — верхние бугорки четверохолмия; 4 — задняя спайка; 5 — пре-тектальная область; 6 — ядро оливы; 7 — глазодвигательный нерв; 8 — ресничный ганглий; 9 — короткий ресничный нерв; 10 — сфинктер радужной оболочки; // — переднее срединное ядро; 12 — медиальная колонка соматических нейронов; 13 — латеральная колонка соматических нейронов; 14 — передне-боковой пучок волокон спинного мозга; /5 — цилиоспинальный центр; 16 — дентальная связка; 17 — вентральные корешки; 18 — соединительная веточка; 19 — первый грудной симпатический ганглий; 20 — нижний шейный симпатический ганглий; 21 — средний шейный симпатический ганглий; 22 — верхний шейный симпатический ганглий; 23 — симпатическое сплетение сонной артерии; 24 — глазная ветвь тройничного нерва; 25 — носо-ресничный нерв; 26 — длинный ресничный нерв; 27 — дилятатор радужной оболочки |
| 22 |
| 18 |

 |
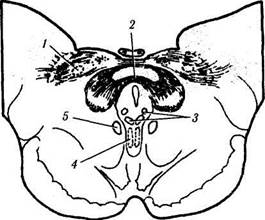 |
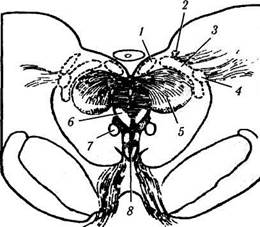
Рис. 4.5.11. Схематическое изображение локализации висцеральных ядер глазодвигательного нерва в дорзальной
части среднего мозга (по Carpenter, Pierson, 1973):
а — отношения переднего срединного ядра, ядра Якубовича— Эдингера—Вестфаля с ядрами претектальной области (/ — ядро оливы; 2 — задняя спайка; 3 — латеральные и медиальные клеточные колонки; 4 — переднее срединное ядро; 5 — ядро Каха-ла). Ядро Якубовича—Эдингера—Вестфаля состоит из двух клеточных групп — латеральной и медиальной клеточных колонок. Переднее срединное ядро расположено непосредственно вент-
ральней и рострально висцеральных клеточных колонок ядра Якубовича—Эдингера—Вестфаля; б — большое претектальное ядро и его отношение с передним срединным ядром (/ — область претектальных ядер; 2 — ядро зрительного тракта; 3 — подчечевицеподобное ядро; 4 —• ядро оливы; 5 — ядро задней спайки; б — ядро Даршкевича; 7 — ядро Кахала; 8 — висцеральное глазодвигательное ядро)
Глава 4. ГОЛОВНОЙ МОЗГ И ГЛАЗ
 в задней спайке, а также в вентральной части сильвиевого водопровода и направляются к «центру сфинктера» как с этой же стороны, так и на противоположную сторону, проходя через среднюю продольную связку (рис. 4.5.7). Число перекрещивающихся аксонов примерно такое же, как и неперекрещивающихся. Из-за симметрического перекреста волокон зрачки обоих глаз обычно одинакового размера. Физиологи предложили модель зрачковой функции, в которой каждый глаз производит сигнал, пропорциональный логарифму интенсивности света, а размер зрачка определяется в среднем мозге взвешенным арифметическим силы двух поступивших сигналов.
в задней спайке, а также в вентральной части сильвиевого водопровода и направляются к «центру сфинктера» как с этой же стороны, так и на противоположную сторону, проходя через среднюю продольную связку (рис. 4.5.7). Число перекрещивающихся аксонов примерно такое же, как и неперекрещивающихся. Из-за симметрического перекреста волокон зрачки обоих глаз обычно одинакового размера. Физиологи предложили модель зрачковой функции, в которой каждый глаз производит сигнал, пропорциональный логарифму интенсивности света, а размер зрачка определяется в среднем мозге взвешенным арифметическим силы двух поступивших сигналов.
«Центр сфинктера» сформирован ядрами Якубовича—Эдингера—Вестфаля глазодвигательного нерва и нейронами претектального ядра [51, 100, 101, 289, 491, 608, 609]. Многие клетки добавочных глазодвигательных ядер, если не их большая часть, обеспечивают механизмы аккомодации. Попытки отличать центры, сужающие и расширяющие зрачок, от «аккомодационных» до настоящего времени полностью успехом не увенчались (см. выше).
Эфферентный путь. Аксоны добавочных глазодвигательных нейронов проходят в составе глазодвигательного нерва (III), располагаясь при этом на его дорзомедиальной поверхности [548]. Отсюда волокна направляются медиально и вниз, поступают в нижнюю ветвь глазодвигательного нерва, с которым и проникают в глазницу (рис. 4.5.10). Большинство волокон в глазодвигательном нерве лежат поверхностно под эпиневрием [301—304].
От нижней ветви глазодвигательного нерва, посредством ветви, идущей к нижней косой мышце, волокна достигают ресничного ганглия (рис. 4.5.2—4.5.5). Эти мякотные преганглио-нарные парасимпатические волокна заканчиваются на теле и дендритах ганглиозных нейронов. Этот парасимпатический узел состоит из двух групп нейронов, меньшая из которых функционально связана с сужением зрачка, а большая — с процессом аккомодации.
Мякотные постганглионарные волокна покидают ресничный ганглий и поступают в глазное яблоко в составе коротких ресничных нервов. Большинство волокон этого пути (90%) направляется к ресничной мышце и только 3—5% к радужной оболочке. Остальные волокна иннервируют кровеносные сосуды и слезную железу. Приведенная рефлекторная дуга обеспечивает рефлекторное сужение зрачка при ярком освещении.








