 2015-07-03
2015-07-03 3763
3763Особенности корневой системы как органа поглощения воды. Д.А. Сабинин, анализируя поглотительную деятельность корневой системы, обратил внимание на то, что скорость передвижения воды в почве крайне мала. За счет диффузии влага в почве передвигается не более чем на 1 см/сут. Уже при поглощении воды со скоростью 1 мг/(ч*см2) быстро наступает иссушение почвы. Следовательно, в почве не вода движется к корню, а корень движется к воде в процессе роста. Н.А. Максимов очень образно писал об этом: «вопреки ходячему представлению, корневая система вовсе не неподвижна закреплена в определенном участке почвы, но все время движется в ней вперед и вперед, словно огромная стая мелких роющих животных, «обсасывая» каждую встречную песчинку и «слизывая» с нее, если так можно выразиться, те тончайшие пленки воды, которые ее одевают».
Это обстоятельство предопределяет специфическую организацию корневой системы – ее большие размеры и сильную разветвленность. Общая поверхность корней обычно превышает поверхность надземных органов в 130-140 раз. Уже у однолетнего сеянца яблони формируется 5-7 порядков ветвления корней общей протяженностью 250 м, а с корневыми волосками около 3 км. У взрослых растений корневая система измеряется десятками километров. Плотность корней древесных и травянистых растений в пахотном слое достигает 0,3-5 см на 1 см3 почвы, тогда как для успешного азотного питания достаточно 01-1 см/см3.
Уникальная возможность корней осваивать новые объемы почвы, иссушив пространства, непосредственно прилегающие к ним, связана с наличием большого количества точек роста (на меристематические ткани приходится 10% массы корня, в стебле – только 1%), высокой скоростью ростовых процессов (1-10см/сут) и положительным гидротропизмом. Благодаря корневым волоскам увеличивается эффективный радиус корня. Корневые волоски играют заметную роль в снабжении растений водой и минеральными веществами, только если их диффузия к поверхности корня затруднена. В водной культуре корневые волоски обычно не образуются.
Следовательно, решающее значение для функционирования корневой системы как поглощающего воду органа является не столько ее общая протяженность или поглощающая поверхность, динамический признак – скорость новообразования и роста корней. У растения ржи, выращенного в искусственных условиях, общая длина всех корней увеличивалась за сутки на 5 км. Однако в биогеоценозе с большой густотой стояния растений размеры корневой системы значительно меньше. Недостаток влаги в почве вызывает торможение роста надземных органов, рост же корневой системы сначала может стимулироваться. Исследование как травянистых, так и древесных растений показали, что водный дефицит и хорошая аэрация почвы стимулируют образование более мощной корневой системы и проникновение ее в более глубокие слои почвенного профиля. Это защитная реакция растений, направленная на добывание воды. при этом надо иметь в виду, что при недостатке воды рост корня ориентирован по градиенту влажности, а не силы тяжести. В засушливых условиях формируется в 3-4 раза большая корневая система, чем во влажных. Во время засухи сначала тормозится рост побегов, а потом уже корней. Усиленное разрастание корневой системы отрицательно сказывается на приросте надземной массы и продуктивности растений. Оптимальное соотношение надземной и подземной массы обеспечивается достаточным водоснабжением и питанием сельскохозяйственных культур и является залогом их высокой продуктивности.
Особенности анатомического строения корня, обуславливающие поглотительную функцию. Известно, что корень делится на четыре зоны (рис.8): деления клеток, растяжения, всасывания (или корневых волосков), и проводящую (или зону опробковения).
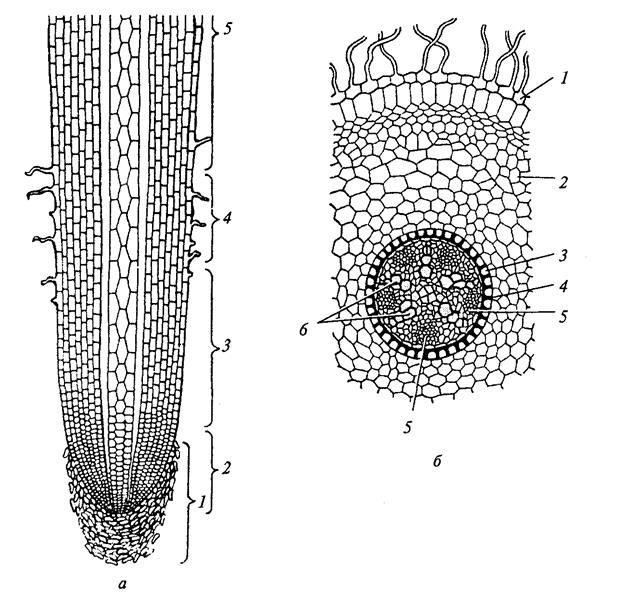
Рис. 8 Строение корня:
а – зоны корня: 1 – корневой чехлик; 2 – зона деления клеток; 3 – зона растяжения клеток; 4 – зона дифференцировки клеток (зона корневых волосков); 5 - зона опробковения (зона проведения); б – поперечный разрез: 1 – корневой волосок; 2 – первичная кора; 3 – эндодерма; 4 – перицикл; 5 – ситовидные трубки; 6 - сосуды
Зона деления клеток, защищенная корневым чехликом, нуждается в небольшом количестве воды, клетки характеризуются корневыми ядрами, большой насыщенностью цитоплазмой, отсутствием вакуолей, первичным строением клеточных оболочек. Их водный потенциал определяется в основном матричным потенциалом, то есть способностью к набуханию коллоидов протоплазмы и клеточных стенок (- 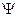 = - m).
= - m).
Поглощение воды интенсивно начинается с зоны растяжения. Здесь идет усиленное новообразование белков цитоплазмы (их содержание в пересчете на клетку возрастает в 1,5 – 2 раза), что усиливает возможности матричного связывания воды. значительное увеличение объема клеток достигается за счет образования крупной центральной вакуоли, которая служит резервуаром осмотически активных веществ. Так появляется второй компонент водного потенциала – осмотический. Одновременно с возрастанием объема наблюдается размягчение и растяжение клеточной оболочки. Благодаря эластичности они не оказывают сопротивления поглощению воды. поэтому водный потенциал определяется суммой матричного и осмотического потенциалов: - = (- m) + (- π) и обеспечивает коллосальную способность поглощать воду. Содержание воды в расчете на клетку возрастает от (1-5)*10-82 в зоне деления до (6-35)*10-82 при растяжении.
Зона корневых волосков является основной поглощающей зоной корня, которая направляет воду в русло дальнего транспорта. Эпиблема покрывает молодые корни. Она состоит из одного слоя клеток. Наружные стенки клеток эпиблемы не имеют ни кутикулы, ни воска, поэтому они хорошо приспособлены для поступления воды. Некоторые клетки эпиблемы вытягиваются и превращаются в корневые волоски. Следовательно, эпиблема как поглощающая ткань неоднородна. Благодаря корневым волоскам поверхность корня, соприкасающаяся с почвой, увеличивается в 10-15 раз. Кроме того, транспортные белки, находящиеся в плазмалемме корневых волосков, активнее, чем в других клетках. Из корневых волосков идет больше плазмодесм в клетки экзодермы. Корневые волоски живут несколько дней и отмирают. Вместо них в верхней части зоны растяжения образуются новые. Постепенно по мере роста корня эпиблема заменяется пробкой. Сильная вакуолизация клеток зоны всасывания (с корневыми волосками и без них), высокая степень развития мембранных структур и ограниченная растяжимость закончивших формирование клеточных стенок выполняют роль механизмов осмотической регуляции транспорта воды, при этом наряду с осмотическими силами большое значение имеет развиваемое гидростатическое давление (Ψр). Поэтому –Ψ = (-Ψπ) + Ψр.
Поглотительная функция проводящей зоны корня из-за опробковения покровных тканей заметно снижена. Однако и опробковевшая часть корня может поглощать воду. Это имеет особое значение для многолетних растений. У растений, обладающих микоризой, последняя выполняет функцию дополнительной поглощающей поверхности, особенно в более старых частях корня.
Механизм поступления воды из почвы в корень. Почти вся поглощаемая растением вода поступает в него через корни. Лишь незначительное количество воды поглощают надземные органы растений. Для поступления воды в корень необходимо, чтобы водный потенциал почвы был более высоким, чем водный потенциал корня. То есть должен быть градиент водного потенциала между почвенным раствором и клетками корня.
Величина водного потенциала почвы определяется выражением:
Ψпоч. = Р – πпоч. – mпоч.,
где Р, πпоч, mпоч – гидростатическое, осмотическое и матричное давление в почве. Величина Ψпоч. Сильно варьирует в зависимости от типа почвы и окружающих условий. После дождя водный потенциал почвы равен нулю, и вода легко поступает в корни. По мере подсыхания почвы её водный потенциал понижается (становится отрицательным). Когда водный потенциал почвы ниже водного потенциала клеток корня, растения завядают. Такую степень увлажнения почвы называют влажностью завядания. Для разных почв она различна: для песка – 1,3 %, для суглинка – 14,5 %. Устойчивое завядание растений часто происходит, когда Ψпоч. падает ниже -1,5 МПа. В этих условиях ни одно растение не может получить воду из почвы. Можно считать, что мезофиты хорошо обеспечены водой, если Ψпоч. не опускается ниже -0,5 МПа. В природных условиях, а также при выращивании отдельных растений без полива в средней климатической зоне Ψпоч. колеблется в диапазоне от -0,5 до -1,2 МПа. В этих условиях вода остается доступной для растений без заметного снижения скорости её поглощения и подавления роста.
Градиент водного потенциала между почвой и клетками корня создается с помощью двух механизмов. Во-первых, благодаря активному поглощению клетками веществ из почвы и, во-вторых, благодаря испарению воды из листьев. В результате этих двух процессов повышается концентрация клеточного сока в клетках корня. Чем меньше насыщенность клеток водой, тем меньше их водный потенциал (более отрицателен). Поступление воды в корень идет в сторону более отрицательного водного потенциала.

