 2015-07-03
2015-07-03 4221
4221Движение воды в радиальном направлении из почвенного раствора в проводящие элементы ксилемы осуществляется как непосредственно через клетки, так и по апопласту тканей корня (рис. 9).
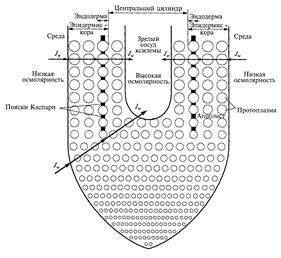
Рис. 9 Транспортная модель корня
(по E. Steudle and C.A. Peterson, 1998)
Апопласт представлен единой системой клеточных стенок и межклеточных полостей (межклетников). Транспорт воды через клетки получил название движение воды от клетки к клетке («cell to cell»). В данном случае вода может идти двумя путями: может пересекать мембраны и идти по непрерывному цитоплазматическому пути через соединяющие соседние клетки плазмодесмы. Единая система, объединяющая с помощью плазмодесм протопласты растительных клеток, называется симпластом. Таким образом, путь движения воды от клетки к клетке подразделяется на трансмембранный и симпластический. В трансмембранном движении воды важную роль играют аквапорины, образующие водные каналы. В итоге, движение воды от клеток ризодермы к сосудам ксилемы корня может осуществляться по трем параллельным направлениям: 1) через клеточные мембраны (трансмембранный путь); 2) по цитоплазме и плазмодесмам (симпластный путь); 3) по клеточным стенкам и межклетникам (апопластный путь). Эти три пути движения воды неавтономны: вода может перемещаться по комбинированным траекториям. Например, пройти некоторый путь по апопласту, затем пересечь плазматическую мембрану и далее передвигаться от клетки к клетке по симпласту.
В настоящее время отсутствуют экспериментальные подходы, которые позволили бы количественно оценить трансмембранный и симпластный потоки воды в корне. Однако имеющиеся данные позволяют предположить, что трансмембранные потоки значительно превосходят симпластные. Из-за отсутствия возможности разделить эти два потока, как правило, пользуются упрощенной транспортной моделью, в которой рассматривают два параллельных пути – путь от клетки к клетке и апопластный путь. Оба потока существуют в растении одновременно. Вклад того и/или иного пути в суммарный поток воды определяется многими факторами: видом растения, его возрастом, условиями выращивания и др.
При апопластном транспорте вода ни разу не пересекает мембраны, а сам апопласт не обладает свойством полупроницаемости. Это означает, что осмотические явления не вовлечены в формирование движущих сил апопластного транспорта воды. В этой связи ток воды через апопласт имеет гидравлическое происхождение, так как движущей силой для него служит преимущественно градиент гидростатического давления (∆Р), и ток воды по апопласту называется гидравлическим. Основным процессом, приводящим к формированию ∆Р, при апопластном (гидравлическом) транспорте является транспирация. При отсутствии водного дефицита в дневное время, когда устьица открыты и интенсивность транспирации высока, поток воды через апопласт в значительной степени превышает поток через клетки. При низкой интенсивности транспирации преобладающим становится движение воды от клетки к клетке.
При движении воды от клетки к клетке, когда вода на своем пути пересекает мембраны, явление осмоса играет определяющую роль. Ток воды в этом случае называется осмотическим, хотя градиент водного потенциала для него включает не только осмотическую, но и гидравлическую составляющую. Градиент водного потенциала между двумя клетками равен:
∆Ψ = ∆Р - δ∆π,
где ∆π, ∆Р – разности тургорного и осмотического давления между двумя клетками;
δ – коэффициент отражения мембран (он стремится к единице, поэтому ∆Ψ = ∆Р - ∆π).
Следует обратить внимание на то, что ∆Р при апопластном токе – это градиент гидростатического давления, создаваемого натяжением жидкости в сосудах ксилемы, а ∆Р при транспорте воды от клетки к клетке – это градиент тургорного давления в клетках корня.
На пути радиального транспорта воды в корне по апопласту расположена эндодерма с поясками Каспари. Основным химическим компонентом поясков Каспари является не суберин, как полагали ранее, а лигнин. Лигнин – это гидрофильное соединение, поэтому он не может препятствовать апопластному пути движения воды. До недавнего времени считали, что движение воды по апопласту в радиальном направлении корня прерывается на уровне эндодермы. Поэтому, чтобы продолжить свое движение в радиальном направлении корня, вода должна перейти в симпласт. Некоторое количество суберина в поясках Каспари может уменьшить гидравлическую проводимость. Таким образом, апопласт эндодермы не оказывает существенного влияния на водный ток, однако является препятствием для движения ионов. Так, на молодых корнях кукурузы было показано, что повреждение эндодермы приводит к выходу ксилемного раствора через повреждение из компартмента с высоким осмотическим давлением (центральный цилиндр) в компартмент с низким давлением (внешняя среда). Эти результаты подтвердили давно предполагавшуюся роль эндодермы как барьера для обратной диффузии ионов из центрального цилиндра в апопласт тканей коры.

