2015-07-21
2015-07-21 639
6391 ) Аминокислоты (и их производные). К ним относят таурин, норадреналин, ДОФАминГАМК, глицин, ацетилхолин, гомоцистеин и некоторые другие (адреналин, серотонин, гистамин, серотонин).
ТАУРИН.
Таурин образуется из аминокислоты цистеина. Сначала происходит окисление серы в SH-группе до остатка серной кислоты (процесс идет в несколько стадий), а затем происходит декарбоксилирование. Таурин - это необычная кислота, в которой нет карбоксильной группы, а имеется остаток серной кислоты:
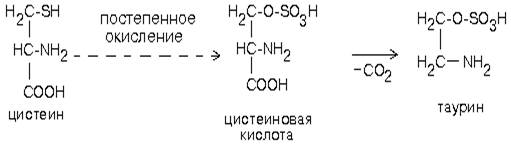
Таурин принимает участие в проведении нервного импульса в процессе зрительного восприятия.
АЦЕТИЛХОЛИН
Для синтеза холина требуются аминокислоты серин, метионин. Этаноламин может быть использован и в готовом виде. Но, как правило, из крови в нервную ткань пступает уже готовый холин. Второй же предшественник этого нейромедиатора - Ацетил-КоА, синтезируется в нервных окончаниях.
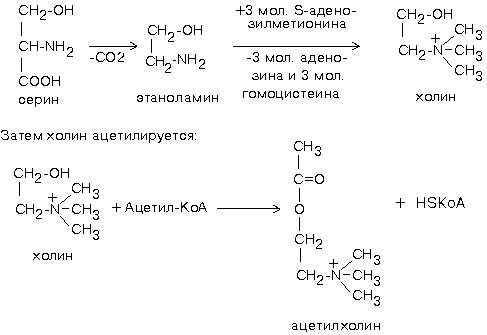
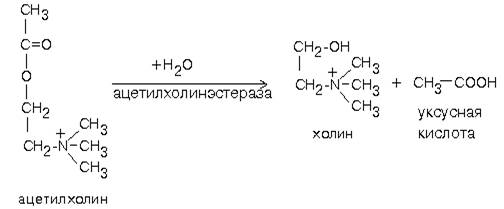
Продукт этой реакции ацетилхолин участвует в синаптической передаче нервного импульса. Он накапливается в синаптических пузырьках, образуя комплексы с отрицательно заряженным белком везикулином. Передача возбуждения с одной клетки на другую осуществляется с помощью специального синаптического механизма.
Синапс - это функциональный контакт специализированных участков плазматических мембран двух возбудимых клеток. Синапс состоит из пресинаптической мембраны, синаптической щели и постинаптической мембраны. Мембраны клеток в месте контакта имеют утолщения в виде бляшек - нервных окончаний. Нервный импульс, достигший нервного окончания, не в состоянии преодолеть возникшее перед ним препятствие - синаптическую щель. После этого электрический сигнал преобразуется в химический. Пресинаптическая мембрана содержит специальные канальные белки, подобные белкам, формирующим натриевый канал в мембране аксона. Они тоже реагируют на мембранный потенциал, изменяя свою конформацию и формируют канал. В результате ионы Са2+ проходят через пресинаптическую мембрану по градиенту концентраций в нервное окончание. Градиент концентраций Са2+ создается работой Са2+-зависимой АТФазы - кальциевым насосом. Повышение концентрации Са2+ внутри нервного окончания вызывает слияние 200-300 имеющихся там везикул, заполненных ацетилхолином, с плазматической мембраной. Далее ацетилхолин секретируется в синаптическую щель путем экзоцитоза. и присоединяется к рецепторным белкам, расположенным на поверхности постсинаптической мембраны.
Ацетилхолиновый рецептор представляет собой трансмембранный олигомерный гликопротеиновый комплекс, состоящий из 6 субъединиц:2-альфа, 2-бета, 1-гамма и 1-дельта. Плотность расположения белков-рецепторов в постсинаптической мембране очень велика - около 20000 молекул на 1 мкм2. Пространственная структура рецептора строго сооответствует конформации медиатора.
При взаимодействии с ацетилхолином белок-рецептор так изменяет свою конформацию, что внутри него формируется натриевый канал. Катионная селективость канала обеспечивается тем, что ворота канала сформированы отрицательно заряженными аминокислотами. Таким образом повышается проницаемость постсинаптической мембраны для натрия и возникает новый импульс (или сокращение мышечного волокна). Деполяризация постсинаптической мембраны выеывает диссоциацию комплекса “ацетилхолин-белок-рецептор” и ацетилхолин освобождается в синаптическую щель. Как только ацетилхолин оказывается в синаптической щели, он за 40мкс подвергается быстрому гидролизу под действием фермента ацетилхолинэстеразы: