 2014-01-31
2014-01-31 806
806Процессы распада нуклеиновых кислот,
В клетках организмов постоянно происходит расщепление полинуклеотидов на олиго- и мононуклеотиды с участием ферментов, называемых нуклеазами. Ранее уже рассматривалось действие различных нуклеаз, участвующих в процессах репарации ДНК, процессинге и сплайсинге РНК-транскриптов, быстрой деградации молекул mРНК. Нуклеазы катализируют гидролиз фосфодиэфирных связей в молекулах РНК и ДНК, а также их ферментов и в зависимости от расщепляемого субстрата эти ферменты называют рибонуклеазами или дезоксирибонуклеазами.
Отдельные нуклеазы очень сильно различаются по специфичности действия и образуемым продуктам. Известны нуклеазы, обладающие невысокой специфичностью. Они способны катализировать гидролиз фосфоэфирных связей в одноцепочечных фрагментах РНК и ДНК с образованием олиго- и мононуклеотидов, имеющих на 5'-конце фосфатные группировки. К таким ферментам относится нуклеаза S1 , выделенная из Aspergillus oryzae.
Неспецифические рибонуклеазы могут действовать на двухцепочечные участки РНК с образованием 5' или 3'-фосфорилированных олигонуклеотидов. Например, рибонуклеаза III Е. coli гидролизует двутяжевые участки в молекулах РНК, образуя в качестве конечных продуктов 3'-фосфорилированные олигонуклеотиды. Неспецифические дезоксирибонуклеазы гидролитически расщепляют двухцепочечные ДНК на олиго- или мононуклеотиды, имеющие фосфатные группировки на 5' или 3'-конце. Например, дезоксирибонуклеаза I из поджелудочной железы животных расщепляет двуспиральные молекулы ДНК на олигонуклеотиды, которые фосфорилированы по 3'-концам.
Специфические нуклеазы осуществляют направленный гидролиз полинуклеотидов, распознавая при своём действии или определённые нуклеотидные радикалы, или специфические последовательности нуклеотидных остатков в молекулах РНК или ДНК. Так, гуаниловая рибонуклеаза Т1 актиномицетов направленно гидролизует фосфодиэфирные связи в одноцепочечных РНК между остатками гуаниловой кислоты и любыми другими нуклеотидными остатками с образованием олигонуклеотидов, имеющих на 3'-конце остатки гуанозин-3- фосфата.
Среди специфических дезоксирибонуклеаз наиболее хорошо изучены рестриктазы, которые рапознают в молекулах ДНК специфические последовательности из 4-6 нуклеотидных остатков. Они расщепляют двухцепочечные молекулы ДНК на строго определённые фрагменты. В качестве примера можно рассмотреть действие рестриктазы Eco RI из клеток E. сoli. Этот фермент распознаёт в двуспиральной ДНК комплементарную нуклеотидную последовательность и гидролитически расщепляет межнуклеотидные связи в положениях, указанных стрелками.
¯
ГААТТЦ
ЦТТААГ
В результате действия этого фермента образуется специфический набор двухцепочечных фрагментов ДНК, содержащих на 5'-концах остатки фосфорной кислоты. В связи с высокой специфичностью действия рестриктазы широко используются для выяснения структуры нуклеиновых кислот.
Под действием набора разных нуклеаз нуклеиновые кислоты гидролизуются до свободных нуклеотидов, которые далее подвергаются распаду с участием гидролитических и окислительных ферментов.
С учасием нуклеотидаз нуклеотиды гидролизуются до нуклеозидов, которые далее под действием нуклеозидфосфорилазы подвергаются фосфоролизу, превращаясь в свободные азотистые основания и фосфорилированные остатки пентозы в виде рибозо-1-фосфата или дизоксирибозо-1-фосфата. Распад нуклеиновых кислот до азотистых оснований может быть представлен в виде следующей схемы.
|
Фосфорилированные пентозы включаются в углеводный обмен, а азотистые основания могут включаться в два типа биохимических процессов – вновь вступать в синтез нуклеотидов или подвергаться дальнейшему распаду.
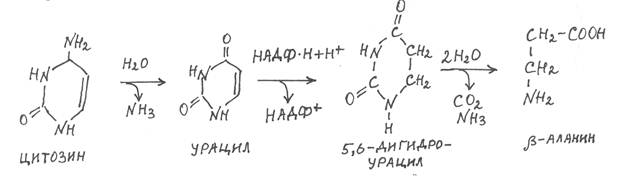
Под действием ферментов фосфорибозилтрансфераз все азотистые основания, кроме тимина, способны взаимодействовать с фосфорибозилпирофосфатом и снова включаться в рибонуклеотиды, которые далее используются для синтеза нуклеиновых кислот. В ходе распада азотистых оснований цитозин в результате гидролитического дезаминирования превращается в урацил, который восстанавливается НАДФ×Н в 5,6-дигидроурацил, а последний подвергается гидролити-ческому расщеплению с образованием b-аланина, СО2 и NH3:
В свою очередь b-аланин может подвергаться окислительному расщеплению и превращаться в малонил-КоА, который при декарбок-силировании даёт ацетил-КоА. А возможные пути метаболизма малонил-КоА и ацетил-КоА нам уже известны.
Тимин в процессе распада подвергается таким же превращениям, как и урацил, только вместо b-аланина образуется b-аминоизобутират, который окислительным путём превращается в метилмалонил-КоА и далее в сукцинил-КоА, а последний включается в дыхательные реакции.
|
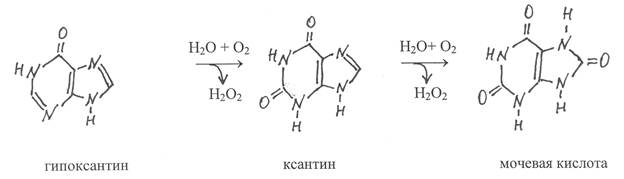
Пуриновые основания аденин и гуанин, подвергаясь гидролитическому дезаминированию, превращаются соответственно в гипоксантин и ксантин. Металлосодержащий флавиновый фермент ксантиноксидаза (1.2.3.2) окисляет гипоксантин в ксантин и далее ксантин в мочевую кислоту. Коферментом в молекуле этого фермента является ФАД, ассоциированный с одним атомом молибдена и четырьмя атомами железа. Всего в димерной молекуле фермента содержится два ФАД, два атома молибдена и восемь атомов железа.
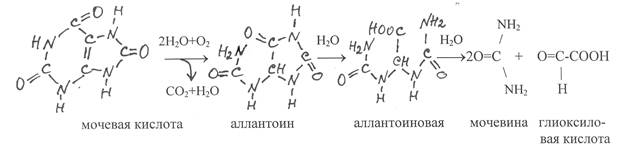
У человека мочевая кислота является конечным продуктом распада пуринов, который выводится из организма. Однако у многих других организмов, включая растения, мочевая кислота окисляется в аллантоин. Эту реакцию катализирует медьсодержащий фермент уратоксидаза (1.7.3.3).
Аллантион в значительном количестве содержится в проростках, молодых побегах и пасоке некоторых растений. С участием фермента аллантоиназы (3.5.2.5) аллантоин гидролизуется с образованием аллантоиновой кислоты, которая также широко распространена в растениях. В клетках растений, плесневых грибов и многих рыб аллантоиновая кислота гидролитически разлагается на мочевину и глиоксиловую кислоту. Эту реакцию катализирует фермент аллантоиказа (3.5.3.4).
Таким образом, пуриновые основания в растительных клетках в результате гидролитических и окислительных реакций так же, как и в орнитиновом цикле, превращаются в мочевину, которая может затем использоваться в метаболизме азотистых веществ по уже известным нам механизмам. Следует отметить, что при высоком содержании в пище человека или кормах животных нуклеиновых кислот в результате диссимиляции пуриновых нуклеотидов образуется много мочевой кислоты, которая у этих организмов является конечным продуктом распада пуриновых оснований. При этом создаётся опасность возникновения мочекаменных заболеваний, вызванных отложением натриевых и кальциевых солей мочевой кислоты.








