 2015-04-30
2015-04-30 3557
3557Структурная организация фибриллярных белков имеет ряд особенностей по сравнению с глобулярными белками. Эти особенности можно проследить на примере кератина, фиброина и коллагена. Кератины существуют в α- и β-конформациях. α-Кератины и фиброин имеют слоисто-складчатую вторичную структуру, однако в кератине цепи параллельны, а в фиброине антипараллельны (см. рис. 3); кроме того, в кератине имеются межцепочечные дисульфидные связи, а у фиброина они отсутствуют. Разрыв дисульфидных связей приводит к разъединению полипептидных цепей в кератинах. Напротив, образование максимального числа дисульфидных связей в кератинах путем воздействия окислителей создает прочную пространственную структуру. Вообще у фибриллярных белков в отличие от глобулярных порой трудно строго разграничить разные уровни организации. Если принять (как для глобулярного белка), что третичная структура должна образовываться путем укладки в пространстве одной полипептидной цепи, а четвертичная - нескольких цепей, то в фибриллярных белках уже при формировании вторичной структуры участвует несколько полипептидных цепей. Типичным примером фибриллярного белка является коллаген, который относится к самым распространенным белкам организма человека (около 1/3 от массы всех белков). Он содержится в тканях, обладающих высокой прочностью и малой растяжимостью (кости, сухожилия, кожа, зубы и т. д.). В коллагене треть аминокислотных остатков приходится на глицин, а около четверти или чуть более — на пролин или гидроксипролин.
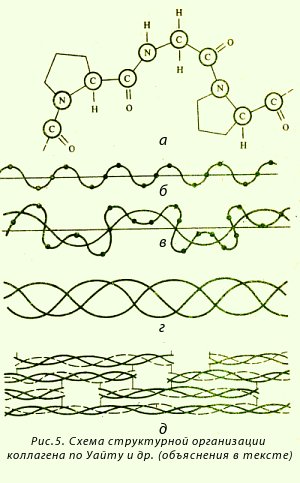
Изолированная полипептидная цепь коллагена (первичная структура) похожа на ломаную линию. Она содержит около 1000 аминокислот и имеет молекулярную массу порядка 105 (рис. 5, а, б). Полипептидная цепь построена из повторяющейся тройки аминокислот (триплет) следующего состава: гли-А-В, где А и В - любые, кроме глицина, аминокислоты (чаше всего пролин и гидроксипролин). Полипептидные цепи коллагена (или α-цепи) при формировании вторичной и третичной структур (рис. 5, в и г) не могут давать типичных α-спиралей, имеющих винтовую симметрию. Этому мешают пролин, гидроксипролин и глицин (антиспиральные аминокислоты). Поэтому три α-цепи образуют как бы скрученные спирали подобно трем нитям, обвивающим цилиндр. Три спиральные α-цепи формируют повторяющуюся структуру коллагена, которая называется тропоколлагеном (рис. 5, г). Тропоколлаген по своей организации является третичной структурой коллагена. Плоские кольца пролина и оксипролина, регулярно чередующиеся вдоль цепи, придают ей жесткость, как и межцепочечные связи между α-цепями тропоколлагена (поэтому коллаген устойчив к растяжению). Тропоколлаген является, по существу, субъединицей фибрилл коллагена. Укладка тропоколлагеновых субъединиц в четвертичную структуру коллагена происходит ступенеобразно (рис. 5, д).
Стабилизация структур коллагена происходит за счет межцепочечных водородных, ионных и ван-дер-ваальсовых связей и небольшого количества ковалентных связей.
α-Цепи коллагена имеют разное химическое строение. Различают α1-цепи разных видов (I, II, III, IV) и α2-цепи. В зависимости от того, какие α1- и α2-цепи участвуют в образовании трехцепочечной спирали тропоколлагена, различают четыре типа коллагена:
· первый тип — две α1 (I) и одна α2-цепи;
· второй тип — три α1 (II)-цепи;
· третий тип — три α1 (III)-цепи;
· четвертый тип — три α1 (IV)-цепи.
Наиболее распространен коллаген первого типа: он содержится в костной ткани, коже, сухожилиях; коллаген второго типа содержится в хрящевой ткани и т. д. В одном виде ткани могут быть разные типы коллагена.
Упорядоченная агрегация коллагеновых структур, их жесткость и инертность обеспечивают высокую прочность коллагеновых волокон. Коллагеновые белки содержат также углеводные компоненты, т. е. являются белок-углеводными комплексами.
Коллаген — внеклеточный белок, который образуется клетками соединительной ткани, входящей во все органы. Поэтому с повреждением коллагена (или нарушением его образования) возникают множественные нарушения опорных функций соединительной ткани органов.
Альфа-полипептидная цепь заканчивается комбинацией аминокислот валина-лейцина, а бета- полипептидная цепь — комбинацией валина-гистидина-лейцина. Альфа- и бета-полипептидные цепи в гемоглобиновой молекуле не размещены линейно, это первичная структура. По причине существования интрамолекулярных сил полипептидные цепи скручиваются в форме типичной для белков альфа-геликсовой спирали (вторичная структура). Сама альфа-геликсовая спираль на каждую альфа- и бета-полипептидную цепь огибается пространственно, образуя сплетения овоидной формы (третичная структура). Отдельные части альфа-геликсовых спиралей полипептидных цепей отмечают латинскими буквами от А до Н. Все четыре третично изогнутые альфа- и бета-полипептидные цепи располагаются пространственно в определенном соотношении — кватернерная структура. Они связаны между собой не настоящими химическими связями, а межмолекулярными силами.
Выяснилось, что у человека имеются три основных типа нормального гемоглобина: эмбриональный — U, фетальный — F и гемоглобин взрослого человека — А. HbU (назван по начальной букве слова uterus) встречается в эмбрионе между 7 и 12 неделями жизни, затем он исчезает и появляется фетальный гемоглобин, который после третьего месяца является основным гемоглобином плода. Вслед за этим появляется постепенно обыкновенный гемоглобин взрослого человека, называемый HbA, по начальной букве английского слова "adult". Количество фетального гемоглобина постепенно уменьшается, так что в момент рождения 80% гемоглобина представляет собой HbA и только 20% — HbF. После рождения фетальный гемоглобин продолжает убывать и к 2 – 3 году жизни составляет всего 1 – 2%. То же количество фетального гемоглобина и у взрослого. Количество HbF, превышающее 2%, считается патологическим для взрослого человека и для детей старше 3 лет.
Кроме нормальных типов гемоглобина, в настоящее время известно свыше 50 его патологических вариантов. Они сначала были названы латинскими буквами. Буква В в обозначениях типов гемоглобина отсутствует, т. к. ею обозначен первоначально HbS.
Гемоглобин (Hb) - хромопротеин, присутствующий в эритроцитах и участвующий в транспорте кислорода к тканям. Гемоглобин взрослых людей называется гемоглобином А (Hb A). Молекулярная масса его составляет около 65000 Да. Молекула Hb А имеет четвертичную структуру и включает четыре субъединицы - полипептидные цепи (обозначаемые α1, α2, β1 и β2, каждая из которых связана с гемом.
Запомните, что гемоглобин относится к аллостерическим белкам, его молекулы могут обратимо переходить из одной конформации в другую. При этом изменяется сродство белка к лигандам. Конформация, обладающая наименьшим сродством к лиганду, называется напряжённой, или Т-конформацией. Конформация, обладающая наибольшим сродством к лиганду, называется релаксированной, или R-конформацией.
R- и Т-конформации молекулы гемоглобина находятся в состоянии динамического равновесия:
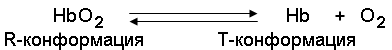
Различные факторы среды могут сдвигать это равновесие в ту или иную сторону. Аллостерическими регуляторами, влияющими на сродство Hb к O2, являются: 1) кислород; 2) концентрация Н+ (рН среды); 3) углекислота (СO2); 4) 2,3-дифосфоглицерат (ДФГ). Присоединение молекулы кислорода к одной из субъединиц гемоглобина способствует переходу напряжённой конформации в релаксированную и повышает сродство к кислороду других субъединиц той же молекулы гемоглобина. Это явление получило название кооперативного эффекта. Сложный характер связывания гемоглобина с кислородом отражает кривая насыщения гемоглобина O2, имеющая S-образную форму (рисунок 3.1).
Повышение содержания СO2, Н+, ДФГ на фоне низкого парциального давления O2 в тканях способствует взаимодействию этих факторов с гемоглобином и переходу R-конформации в Т-конформацию. Это приводит к смещению равновесия в уравнении (1) вправо. Выделившийся O2 поступает в ткани.
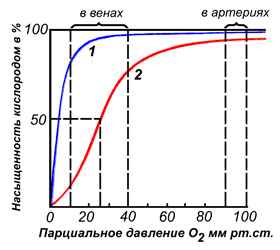
Рисунок 3.1. Кривые насыщения миоглобина (1) и гемоглобина (2) кислородом.
Молекулярные формы гемоглобина отличаются друг от друга строением полипептидных цепей. Примером такой разновидности гемоглобина, существующей в физиологических условиях, является фетальный гемоглобин (HbF), присутствующий в крови в эмбриональной стадии развития человека. В отличие от HbA, его молекула содержит 2 α- и 2 γ-цепи (то есть β-цепи заменены на γ-цепи). Такой гемоглобин обладает более высоким сродством к кислороду. Именно это позволяет эмбриону получать кислород из крови матери через плаценту. Вскоре после рождения HbF в крови ребёнка замещается на HbA.
В качестве примера аномального или патологического гемоглобина можно привести уже упоминавшийся (см. 2.4.) гемоглобин S, обнаруженный у больных серповидно-клеточной анемией. Как вам уже известно, он отличается от гемоглобина А заменой в β-цепях глутамата на валин. Эта аминокислотная замена вызывает снижение растворимости HbS в воде и уменьшение его сродства к O2.
3. Найти самой
4. Определение общего белка в сыворотке крови по биуретовой реакции
Определение общего белка по биуретовой реакции является на сегодняшний день самым распространенным методом определения общего белка в сыворотке крови. Метод относительно дешев, прост, обладает хорошей воспроизводимостью и специфичностью, использование его позволяет выполнять исследование как на анализаторах (автоматических и полуавтоматических), так и на обычном фотометре.








