 2015-04-30
2015-04-30 3727
3727Связь миллиардов нейронов мозга осуществляется посредством медиаторов — ацетилхолин и норадреналин. Содержащие их нервы называют соответственно холинергическими и адренергическими. В соответствии с этим все эфферентные системы делят на холинорецепторы и адренорецепторы.
К другим медиаторам относятся дофамин, адреналин, серотонин, октопамин, гистамин, ГАМК и др.
Обширная группа холинорецепторов весьма неоднородна как в структурном, так и в функциональном отношении. Объединяют их медиатор — ацетилхолин — и общая схема строения синапса.
 |
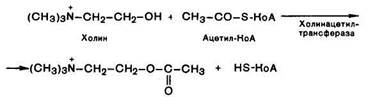
Ацетилхолин представляет собой сложный эфир уксусной кислоты и холина. Он синтезируется в нервной клетке из холина и активной формы ацетата — ацетилкоэнзима А при помощи специального фермента холинацетилтрансферазы (холйнацетилазы):
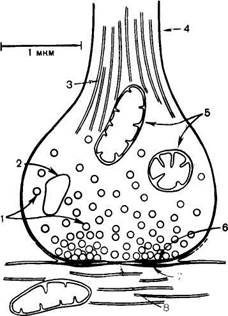
| Схематическое изображение синапса (по Мецлеру). 1 — синаптические пузырьки; 2 — лизосома; 3 — микрофибриллы (нейрофибриллы); 4 - аксон; 5 - митохондрии; 6 — пресинаптическое утолщение мембраны; 7 - постсинаптическое утолщение мембраны; 8 — синаптическая щель (около 20 нм). |
Синапс можно представить себе как узкое пространство (щель), ограниченное с одной стороны пресинаптической, с другой — постсинаптической мембраной. Пресинаптическая мембрана состоит из внутреннего слоя, принадлежащего цитоплазме нервного окончания, и наружного слоя, образованного нейроглией. Мембрана в некоторых местах утолщена и уплотнена, в других — истончена и имеет отверстия, с помощью которых цитоплазма аксона может сообщаться с синаптическим пространством. Постсинаптическая мембрана менее плотная, не имеет отверстий.
В синаптических нервных окончаниях имеются пузырьки (везикулы) диаметром 30—80 нм, которые содержат нейромедиаторы. В холинергических синапсах каждый пузырек содержит молекулы ацетилхолина. При возбуждении высвобождение медиатора происходит «квантами», т. е. путем полного опорожнения каждого отдельного пузырька. Выделенный в синаптическую щель ацетилхолин вступает во взаимодействие с белком-хеморецептором, входящим в состав постсинаптической мембраны. В результате изменяется проницаемость мембраны. Взаимодействие между рецептором и медиатором запускает ряд реакций, заставляющих постсинаптическую нервную клетку или эффекторную клетку выполнять свою специфическую функцию. После выделения медиатора должна наступить фаза его быстрой инактивации или удаления, чтобы подготовить синапс к восприятию нового импульса.
В адренорецепторах существует два вида рецепторов для норадреналина: α- и β-адренергические рецепторы.
β-Адренергические рецепторы включают эфферентную клетку с помощью аденозин-3', 5'-монофосфата, или цАМФ — универсального «второго посредника» между гормонами и различными функциями клеток, на которые воздействуют гормоны.
Как только β-адренергический рецептор (расположенный на наружной поверхности мембраны эффекторной клетки) начнет взаимодействовать с норадреналином, на внутренней поверхности клеточной мембраны активируется фермент аденилатциклаза. Затем в клетке аденилатциклаза превращает АТФ в цАМФ; последний в свою очередь способен оказывать влияние на метаболизм клетки. Известно, что в метаболизме катехоламиновых медиаторов особая роль принадлежит ферменту моноаминоксидазе (МАО). Этот фермент удаляет аминогруппу (-NH2) у норадреналина, серотонйна, дофамина и адреналина, тем самым инактивируя упомянутые медиаторы.
Адренергическая и холинергическая системы головного мозга тесно взаимодействуют с другими системами мозга, в частности использующими серотонин в качестве медиатора. В основном серотонинсодержащие нейроны сосредоточены в ядрах мозгового ствола. Нейромедиаторная роль серотонина осуществляется в результате взаимодействия серотонина со специфическими серотонинергическими рецепторами.
Важным нейромедиатором, выполняющим тормозные функции, является ГАМК, количество которой в головном мозге во много раз выше, чем других нейромедиаторов.








