 2015-05-13
2015-05-13 2556
2556В палеогене начинается новый этап в развитии органического мира. На рубеже мезозоя и кайнозоя вымерли многие характерные для мезозоя морские и наземные животные, а сохранившиеся в значительной мере утратили свое значение. К этому времени полностью прекращают свое существование аммониты, белемниты, рудисты, иноцерамы, а также ихтиозавры, плезиозавры, динозавры и ряд других мезозойских групп пресмыкающихся. Их место занимают бурно развивающиеся млекопитающие.
На смену мезозойским пришли новые роды и семейства двустворчатых и брюхоногих моллюсков, а также костистых рыб. Резкий толчок в развитии испытала покрытосеменная растительность, которая начала доминировать. Исключительно большое развитие получили фораминиферы. Важнейшими среди планктонных форм были роды Globigerina, Subbotina, Globorotalis, Acarinina, Hantkenina, а среди бентосных крупных
фораминифер – Nummulites, Assilina, Operculina, Discocyclina. Все они очень быстро эволюционировали и особенно широкое развитие получили в эоцене, но уже в начале олигоцена вымерли. Значительным распространением пользовались двустворчатые и брюхоногие моллюски. В отличие от других моллюсков, они обитали не только в морях с нормальной соленостью, но и в солоноватоводных пресных бассейнах. Некоторые гастроподы освоили приморские низменности. Родовые видовые комплексы моллюсков служат основой стратиграфического расчленения палеогена. Характерными и важными являлись Pecten, Cucullaea, Arca, Chlamys, Grassatella, Ostrea, Cardium, а среди гастропод – Turritella, Cerithium, Pleurotoma, Trochus, Natica (рис. 13.1).
Из других беспозвоночных широко были распространены неправильные морские ежи, мшанки, губки, членистоногие, кораллы. Теплые моря с нормальной соленостью были населены ежами Echinocorys, Clypeaster, Cjnjclypeus. Шестилучевые кораллы вместе с остатками водорослей участвовали в строении рифовых массивов. Из членистоногих известны остракоды, среди которых распространены Trachyleberis, Cytheridea, Koczoloncha, меньше – десятиногие ракообразные, в частности пресноводные и морские раки и креветки. Роль мшанок и брахиопод в сравнении существенно сократилась.
В последние годы установлено, что среди палеогеновых организмов большое значение имели представители наннопланктона. Расцвет золотистых микроскопических водорослей был в эоцене. Наряду с ними породообразующую роль играли диатомовые и кремнистые жгутиковые. В морях обитали морские позвоночные, среди которых преобладали костистые. Кроме них известны хрящевые – акуловые и скаты. Появились древние представители китов, сирены, дельфины – млекопитающие, освоившие морские просторы.
Существенные изменения произошли в наземной фауне палеогена. Пресмыкающиеся, некогда господствующие на земной поверхности, уступили место млекопитающим. Из рептилий продолжали существовать крокодилы, ящерицы, змеи. Земноводные представлены гигантскими саламандрами, лягушками и жабами. В воздухе доминировали птицы.
Млекопитающие палеогена были разнообразными. Уже в эоцене широко распространились основные отряды кайнозойских грызунов, хищников, непарно- и парнокопытных. В это же время возникли насекомоядные и зайцеобразные. Самыми крупными млекопитающими в олигоцене были представители древних носорогов – гигантские безрогие индрикотерии (рис. 13.2). Основным местом их обитания были саванны. В начале неогена индрикотерии вымерли. В олигоцене существовали примитивные свиньи, верблюды,


олени. Уже в начале палеогена появились лемуры – наиболее примитивная группа среди
приматов, называемая полуобезьянами. Только в конце эоцена возникли настоящие обезьяны – антропоиды. В это же время появились крупные хоботные и сумчатые. Среди последних известны как хищные, так и растительноядные формы.
Среди наземной флоры продолжалось развитие покрытосеменных. Многие регионы покрывались густыми тропическими и субтропическими лесами. В их составе ведущая роль принадлежала пальмам, магнолиям, миртам, фикусам, гигантским секвойям, араукариевым и кипарисовым. Наряду с ними в более умеренном климате росли широколиственные и мелколистные – дуб, бук, каштан, тополь, береза и другие.
Органический мир неогена имеет типичный кайнозойский облик, который к концу периода приобретает современные черты (рис. 13.3). В морях неогена продолжали развитие те же группы организмов, что и в палеогене. Преобладали простейшие, двустворчатые и брюхоногие моллюски, остракоды. Все они имеют важное стратиграфическое значение.

Реже, чем в более древних отложениях, обнаруживаются остатки мшанок, губок, кораллов, брахиопод, иглокожих, рыб и водных млекопитающих.В составе фораминиферовой фауны произошли крупные изменения. Вымерли нуммулиты, ассилины, дискоциклины, но очень обильной стала фауна планктонных фораминифер. Среди бентосных ассоциаций господство принадлежало двустворчатым и брюхоногим моллюскам, которые в неогене достигли исключительного многообразия. В морях с нормальной соленостью обитали Nukula, Fissurella, Cardita, Venus, Turritella, Cerithium, Conus. В замкнутых опресненных бассейнах, в частности в бассейне Паратетиса, известен совершенно иной комплекс моллюсков: Mactra, Congeria, Ervillia, Mytilus, Dreissena, Didacna. Большим развитием пользуются пресноводные и наземные формы – Unio, Planorbis, Melanopsis, Helix, Viviparis. В конце неогена состав моллюсков близок к современному.
В морях с нормальной соленостью обитали кораллы, но ареал их распространения стал сокращаться. Границы развития рифов постепенно смещались в сторону экватора. В морях обитали губки, радиолярии, иглокожие, мшанки и брахиоподы. В конце неогена их облик приобрел современный вид.
Среди водных позвоночных господствовали костистые рыбы. Их остатки встречаются как в нормально-морских, солоноватоводных, так и пресноводных бассейнах. Много остатков хрящевых рыб. В начале неогена появились ластоногие, тюлени и моржи. Моря этого периода характеризовались богатством одноклеточных водорослей – диатомовых и золотистых (кокколитофориды). Последние имеют важное стратиграфическое значение.
Сильно меняется состав организмов суши (рис. 13.4). Особенно глубокие изменения претерпела фауна млекопитающих, приспособившихся к обитанию в густых лесах, лесостепных, степных и полупустынных районах. Возникли и широко распространились современные семейства и роды хищных, копытных и хоботных. Так в миоцене появились медведи, гиены, куницы, собаки, барсуки, мастодонты, носороги, быки, овцы, а в плиоцене – ласки, росомахи, гиппопотамы, олени, гиппарионы (трехпалые) и настоящие лошади.
В Северной Америке в начале миоцена не было хоботных. Беднее была фауна хищников, но копытные развивались быстрее. В конце миоцена происходит широкий обмен млекопитающими, что свидетельствует о соединении Азии с Северной Америкой. Млекопитающие Южной Америки в неогене находились на более низкой ступени развития, чем в Северной Америке. В первой в миоцене господствовали грызуны и сумчатые, отсутствовали хищные и хоботные. Лишь в середине плиоцена началось интенсивное проникновение на юг североамериканских форм. Наиболее примитивной была фауна млекопитающих Австралии, состоящая из сумчатых и однопроходных. Это объясняется ранней изоляцией Австралии от других материков. Зоогеографические провинции неогена в общих чертах сходны с современными, но отличались очертаниями своих границ и размерами. В связи с резко выраженной изоляцией неогеновых бассейнов их фауна образует комплексы, имеющие чисто местное (эндемичное) значение.
Приматы в неогене обитали не только в лесах, но и стали осваивать открытые пространства. В верхнемиоценовых отложениях Европы обнаружены остатки обезьян – дриопитеков, напоминающих современных шимпанзе. Древнейшим представителем гоминид считается рамапитек, остатки которого обнаружены в отложениях верхнего миоцена Индии и Кении. Много ископаемых остатков гоминид описаны из отложений верхнего плиоцена Восточной и Южной Африки. Все они принадлежат разновидностям австралопитека – собственно австралопитеку, парантропу и зинджантропу. Все перечисленные остатки по строению ближе к скелету людей, чем любая из современных человекообразных обезьян.
Хотя наземная флора неогена по своему составу близка к палеогеновой, но в составе первой появились ассоциации, сильно напоминающие современные. В связи с развитием похолодания состав флоры стал более дифференцированным. Происходит постепенное оттеснение в сторону экватора теплолюбивых форм – пальм, миртовых, лавровых. В умеренных широтах большое развитие получили тополь, березе, ива, клен, грецкий орех, ель, сосна, пихта и другие деревья. Появились и широко распространились в течение неогена лесостепные, степные, таёжные и тундровые ассоциации растительности.
Животный и растительный мир четвертичного периода близок к современному (рис. 13.5). Изменения, происходившие в составе и расселении животных и растений, были связаны с изменениями природной среды, вызванными чередующимися похолоданиями и потеплениями. Во время оледенений холоднолюбивые формы мигрировали в сторону экватора и во внеледниковой зоне наряду с представителями южной фауны и флоры распространены формы, жившие в приполярных широтах. Во время межоледенений миграция происходила в обратном направлении. Сильно расширялись области распространения субтропической, тропической и умеренно-теплой фауны и флоры, а в отдельные, наиболее теплые отрезки времени, исчезали представители тундровых ассоциаций.
Характерной особенностью развития флоры в течение плейстоцена было её неоднократное изменение и приспособление к похолоданиям в умеренных и субтропических широтах, сопровождавшееся обеднением видового и родового состава, исчезновением вечнозеленых и широколиственных форм и расширением ареалов травянистых сообществ.
В животном мире заметные изменения происходили в составе млекопитающих, особенно хоботных и копытных, населявших Северное полушарие. В плейстоцене в результате многократных смен похолоданий и потеплений вымирали многие теплолюбивые животные и появились приспособленные к жизни в суровых условиях. Особенно усилились процессы вымирания и миграции в эпоху максимального днепровского оледенения.
В конце плиоцена и в самом начале четвертичного периода на юге Восточной Европы обитало много теплолюбивых форм, среди которых были мастодонты, южные слоны, слон Громова, гиппарион, саблезубый тигр, этрусский носорог и другие. В это время на юге Западной Европы кроме перечисленных животных жили гиппопотамы и страусы. Однако в течение раннего плейстоцена, задолго до начала максимального оледенения, состав животного мира начал меняться. Особенно значительные изменения произошли в эпоху днепровского оледенения, когда теплолюбивые животные перемещалшись далеко к югу. В этом же направлении смещались границы распространения растительных ассоциаций.
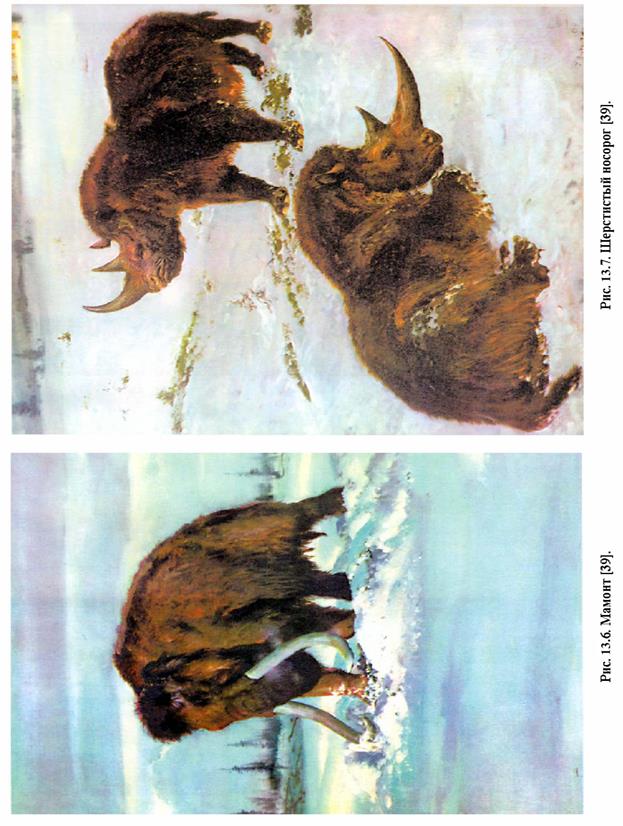
В днепровскую ледниковую эпоху на равнинах Европы южнее границы ледника в перигляциальных областях наряду с оленями, волками, лисицами, бурыми медведями впервые появились такие животные, как мамонт (рис. 13.6), шерстистый носорог, (рис. 13.7), мускусный овцебык, северный олень, белые куропатки, лемминги. Все они были обитателями холодных областей. Из-за резкого и сильного похолодания стали вымирать ранее жившие здесь теплолюбивые организмы, в том числе гигантские носороги, разнообразные эласмотерии, носорог Мерка, пещерные львы, медведи и гиены. Холод в горных областях Кавказа, Кавказа, Крыма, Карпат, Альп, Пиренеев способствовал переселению животных с высокогорий в долины. В Крыму появились белая куропатка, песец, заяц-беляк, северный олень, полярный жаворонок.
Мамонты и шерстистые носороги стали обитать на юге Европы. Они заселили всю Западную Европу и Восточную Сибирь, проникли в Северную Америку. Изоляция способствовала сохранению прежней фауны в Австралии, Южной Америке, Центральной и Южной Африке. Однако периодически наступающие похолодания приводили к существенному обеднению фауны. В конце плейстоцена – начале голоцена вымерли хорошо приспособившиеся к условиям сурового климата мамонты, шерстистые носороги, большерогие олени. В Северной Америке исчезли некоторые млекопитающие и среди них мастодонты и ленивцы.
Наиболее важное событие четвертичного периода – стремительное развитие человека. Полученные в последнее время материалы дали возможность разрешить некоторые проблемы времени и места зарождения древнего человека. Долгое время исследователи


считали, что прародиной человека была Азия. Новейшие открытия показали, что наиболее вероятным местом появления древнего человека была Восточная Африка.
Предковой формой ископаемых людей были австралопитеки, принадлежавшие семейству гоминид (рис. 13.8). Они встречены только на Африканском континенте и впервые появились в Восточной Африке около пяти млн лет назад. Все известные находки австралопитеков сосредоточены в пределах системы Великих Африканских грабенов – крупной рифтовой системы, состоящей из цепи впадин, осложненных поднятиями и занятых озерами. В течение плиоцена здесь господствовал теплый влажный климат и неоднократно происходили извержения вулканов, которые сопровождали рост рифтовой системы.
Природные условия здесь благоприятствовали пышному развитию органического мира, в котором немаловажная роль принадлежала гоминидам. Австралопитеки постепенно освоили прямое хождение и стали всеядными, а 2,6 млн лет назад начали применять примитивно обработанные гальки в качестве орудий. В процессе эволюции среди австралопитеков появились формы, приближающие их к другому, более совершенному виду человека, получившему название Homo habilis – человек умелый.
Следующим звеном в развитии гоминид являются архантропы, которых многие исследователи считают предками ископаемых людей. Они появились также в Восточной Африке, но уже около 1,4-1,2 млн назад и оттуда мигрировали на другие континенты. Наиболее широко известными представителями архантропов являются питекантропы (рис. 13.9). Время их существования носит название раннего палеолита. Находки остатков питекантропов сделаны на острове Ява, в Китае, Венгрии и Германии. Возле остатков питекантропов в Восточной Африке обнаружены каменные орудия в виде ручных рубил. Обработанные камни и следы костров найдены в пещере вместе с остатками синантропов недалеко от г. Пекина. Находки остатков питекантропов в Восточной Африке самые древние, а все остальные имеют более молодой возраст. Скорее всего, питекантропы развивались вплоть до миндельского времени, т.е. до 350-400 тысяч лет назад.
В ряде районов обнаружены остатки ископаемого человека, обладавшего переходными признаками от архантропов к палеоантропам. Судя по остаткам в пещере Азых в Азейбарджане и в районе Арго во Франции, люди широко использовали ручные рубила и применяли огонь. По степени обработки каменных изделий археологи выделяют ашельскую культуру. Палеоантропы или неандертальцы (рис. 13.10) появились около 350 тысяч лет тому назад и существовали до середины вюрма, т.е. около 35 тысяч лет назад. Они обитали на значительно большей территории, чем архантропы. Остатки неандертальцев часто обнаруживают в умеренных и южных широтах Европы. По степени обработки орудий эта археологическая культура стоит выше ашельской и носит название мустьерской.
Представители мустьерской культуры обитали в примитивных жилищах, построенных из деревьев и костей на открытых пространствах. Погребения носителей мустьерской культуры сохранили следы ритуальных действий. Время существования палеоантропов называют палеолитом. Неоантропы (homo sapiens) – люди современного типа – сменили палеоантропов в середине вюрма, т.е. около 35-40 тысяч лет назад во время позднего палеолита. Неоантропы жили в южных и средних широтах Евразии, но иногда появлялись в северных районах. Они проникли в Америку, Австралию и на многочисленные острова Тихого, Атлантического и Индийского океанов.
Уже первые неоантропы (кроманьонцы) мало отличались от современных людей (рис. 13.11). Они быстро и хорошо приспособились к жизни как в жарких, так и холодных условиях. Неоантропы достигли значительных успехов в технике обработки камня. У них появились костяные изделия, предметы изобразительного искусства, в частности наскальные рисунки, статуэтки, гравировка по кости, примитивные музыкальные инструменты, украшения, предметы культа. Люди, жившие в суровых климатических условиях и при периодически наступавших похолоданиях, стали использовать меховую одежду.
Позднее началось разделение сообществ людей по видам добычи продуктов питания. Люди, занимавшиеся в основном охотой, стали объединяться в специализированные племена. Они строили жилища из деревьев и костей, прикрывали их ветвями и звериными шкурами. Уклад жизни, бытовые и хозяйственные элементы у охотничьих племен были иными, чем у тех, что занимались рыбной ловлей или собирательством плодов и растений.
Около 10000 лет назад палеолит сменился мезолитом. Его начало совпадает с границей плейстоцена и голоцена. В это время началось энергичное отступание материковых льдов. Климатический оптимум способствовал расширению благоприятных для жизни людей территорий. Люди приобрели возможность осваивать приполярные районы. В мезолите хозяйственный уклад племен ещё больше усложнился. Появился лук и стрелы,

люди занялись одомашниванием животных, возникли первые скотоводческие и земледельческие племена.
Около 6 тысяч лет назад, а в некоторых регионах и раньше, произошел переход от мезолита к неолиту, т.е. к позднему каменному веку. В это время люди научились обжигать глину и появились предметы из керамики. В неолите бродячий кочевой образ жизни сменился оседлым. Возникли земледельческие культуры, а охотничьи и рыболовные хозяйства сменились земледельческими и скотоводческими. Около 5 тысяч лет назад появились первые бронзовые изделия, 3 – 2,5 тысяч лет назад началась эпоха железа.
Ввиду того, что костные остатки ископаемых людей встречаются довольно редко и к тому же дают возможность охарактеризовать только более или менее крупные стратиграфические рубежи в рамках четвертичного периода, для стратиграфических целей большое значение имеют следы различных культур каменного века. Они совместно с остатками животных и растений, а также палеогеографическими реконструкциями дают возможность разработать более детальные корреляционные схемы.








