 2015-05-26
2015-05-26 4568
4568 ФИЗИОЛОГИЯ РАЗМНОЖЕНИЯ РАСТЕНИЙ
ФИЗИОЛОГИЯ РАЗМНОЖЕНИЯ РАСТЕНИЙ
«Нельзя не заметить, что биология размножения растения весьма слабо разработана экспериментально. До настоящего времени мы вынуждены довольствоваться главным образом описательной стороной весьма многочисленных и разнообразных явлений, стоящих в прямом или косвенном отношении к процессу размножения».
В. Н. Любименко
 В предыдущей главе были рассмотрены все этапы онтогенеза высших растений, за исключением этапа зрелости и размножения. Молодые растения растут и развиваются, создавая достаточно мощную вегетативную массу и запас питательных веществ, для того чтобы, перейдя к этапу размножения, иметь возможность формировать органы размножения, питающиеся гетеротрофно.
В предыдущей главе были рассмотрены все этапы онтогенеза высших растений, за исключением этапа зрелости и размножения. Молодые растения растут и развиваются, создавая достаточно мощную вегетативную массу и запас питательных веществ, для того чтобы, перейдя к этапу размножения, иметь возможность формировать органы размножения, питающиеся гетеротрофно.
Размножение растений — это физиологический процесс воспроизведения себе подобных организмов, обеспечивающий непрерывность существования вида и расселение его представителей в окружающем пространстве.
У растений выделяют три типа размножения: половое, бесполое и вегетативное. При половом размножении новый организм появляется в результате слияния двух половых клеток — гамет. Половой процесс описан для всех высших и низших растений. Бесполое размножение характерно для споровых растений, которые почти всегда имеют более или менее четко выраженное чередование двух поколений — бесполого ди-плоидного (спорофит) и полового гаплоидного (гаметофит). При бесполом размножении новый организм развивается из спор, образующихся на спорофите. Вегетативным размножением называется воспроизведение растений из вегетативных частей растения, т. е. из кусочков таллома или из отдельных клеток у низших растений, из органов или их частей — у высших.
 По мнению В. В. Скрипчинского (1977, 1985), эволюция способов размножения у растений происходила следующим образом. На первом этапе из форм, размножающихся простым делением клеток, возникли спорообразующие, а позже — способные к половому размножению организмы. В результате объединения бесполого и полового типов размножения появились виды с чередованием поколений. У наиболее высокоорганизованных водорослей одно поколение стало доминирующим, наиболее приспособленным к условиям существования. У наземных растений эта тенденция выражена еще более четко: у мхов спорофит вообще не способен к самостоятельному существованию и живет за счет гаметофита. У остальных высших растений, напротив, редукции подвергся гаметофит: если у папоротникообразных он существует еще самостоятельно в виде заростков, то у цветковых растений микрогаметофит редуцирован до 2 — 3 клеток, а макрогаметофит на ранней фазе представляет собой восьмиядерный зародышевый мешок.
По мнению В. В. Скрипчинского (1977, 1985), эволюция способов размножения у растений происходила следующим образом. На первом этапе из форм, размножающихся простым делением клеток, возникли спорообразующие, а позже — способные к половому размножению организмы. В результате объединения бесполого и полового типов размножения появились виды с чередованием поколений. У наиболее высокоорганизованных водорослей одно поколение стало доминирующим, наиболее приспособленным к условиям существования. У наземных растений эта тенденция выражена еще более четко: у мхов спорофит вообще не способен к самостоятельному существованию и живет за счет гаметофита. У остальных высших растений, напротив, редукции подвергся гаметофит: если у папоротникообразных он существует еще самостоятельно в виде заростков, то у цветковых растений микрогаметофит редуцирован до 2 — 3 клеток, а макрогаметофит на ранней фазе представляет собой восьмиядерный зародышевый мешок.
В анатомо-морфологическом плане процессы размножения у многих групп растений изучены довольно подробно, однако их физиологическая основа выяснена далеко недостаточно.
Переход цветковых растений от вегетативного развития и роста к генеративному развитию — сложный многофазный процесс. При достижении зрелого возраста и инициации цветения вегетативные почки преобразуются во флоральные, в которых вместо примордиев листьев закладываются зачатки органов цветка, осуществляется их формирование и рост, вслед за опылением и оплодотворением образуются семена и плоды, распространяющиеся ветром, водой, животными по окружающей территории.
Инициация перехода к зацветанию включает в себя две фазы: индукцию и эвокацию.
Индукция. Эта фаза осуществляется под действием экологических факторов — температуры (яровизация) и чередования дня и ночи (фотопериодизм) — или эндогенных факторов, обусловленных возрастом растения. Яровизация — процесс, протекающий в озимых формах однолетних и двулетних растений под действием низких положительных температур определенной длительности, способствующий последующему ускорению развития этих растений. Фотопериодизм — реакция растений на суточный ритм освещения, т. е. на соотношение длины дня и ночи (фотопериоды}, выражающаяся в изменении процессов роста и развития. Одно из основных проявлений этой реакции — фотопериодическая индукция зацветания. Оба фактора — температурный и световой — могут действовать последовательно, например у озимых злаков (рожь, пшеница). Температурная и фотопериодическая регуляции служат приспособлением растений к условиям существования, так как обусловливают благоприятные сроки для перехода к зацветанию. В ходе фотопериодической индукции в листьях образуется стимулятор цветения, который транспортируется в вегетативные почки побегов, где включает вторую фазу инициации — эвокацию.
Яровизация. Растения, нуждающиеся в яровизации, называют озимыми, а развивающиеся без нее — яровыми. Виды без яровизации не зацветают (качественная реакция), у других при воздействии низкими температурами лишь укорачивается время перехода к цветению (количественная реакция). У растений с количественной реакцией яровизация может проходить в наклюнувшихся семенах. Растения с качественной реакцией (двулетние и многолетние) должны достичь определенных размеров до ее начала.
Яровизация обычно продолжается 1—3 месяца. Наиболее эффективны температуры от 0 до 7 °С. Для теплолюбивых растений оптимальны более высокие температуры порядка 10-13°С. У некоторых видов, например у озимой ржи и капусты, периодическое прерывание низкотемпературного воздействия снижает и даже устраняет эффект яровизации. Необходимое условие яровизации — наличие делящихся клеток. Она протекает в зародыше или в апикальных меристемах стебля и в молодых листьях. Яровизация успешно осуществляется в изолированных верхушках стеблей, например у моркови и капусты. Изменения, возникшие в период яровизации, передаются только путем деления клеток.
В ряде опытов показано, что непрерывный свет способствует переходу растений от озимого поведения к типичному яровому. После яровизации большинству растений необходимы длиннодневные фотопериоды. У других растений яровизация ослабляет или даже устраняет потребность в последующем фотопериодическом воздействии.
Физиолого-биохимическая сущность яровизации до сих пор не ясна. У некоторых видов в ходе яровизации или после ее окончания увеличивается содержание гиббереллинов в тканях.
Фотопериодизм. Понятия фотопериод и фотопериодизм были введены в науку американскими физиологами растений У. Гарнером и Г. А. Аллардом (1920—1923). В зависимости от реакции на длину дня, ускоряющей зацветание, растения делятся на длиннодневные (ДДР), короткодневные (КДР), растения, нуждающиеся в чередовании разных фотопериодов,— длинно-короткодневные (ДКДР) и коротко-длиннодневные (КДДР), а также на нейтральные по отношению к длине дня растения (НДР). К ДДР относятся, в частности, хлебные злаки, многие крестоцветные, укроп и др., к КДР — рис, соя, дурнишник, конопля, к НДР — гречиха, горох и т. д. ДДР распространены в основном в умеренных и приполярных широтах, КДР — в субтропиках.
Отнесение растений к той или иной фотопериодической группе не связано с конкретной оптимальной длиной дня, а дает представление о том, ускоряется ли переход к цветению при увеличении или уменьшении длительности освещения в каждом фотопериоде. Среди растений есть виды и разновидности как с качественным, так и с количественным типами фотопериодических реакций. Интенсивность освещения, температура и другие факторы могут менять характер фотопериодических ответов растений. Для КДР с качественной реакцией решающим фактором служит длина темнового периода. Кратковременное освещение этих растений (1 мин) в середине темнового периода препятствует их переходу к цветению. Прерывание светового периода темнотой не влияет на сроки цветения. ДДР не нуждаются в периодах темноты и зацветают на непрерывном свету.
Фотопериодические воздействия воспринимаются главным образом листьями, а не апексами побега. У большинства растений максимальной чувствительностью к фотопериоду обладают листья, только что прекратившие рост. Для дурнишника, зацветание которого индуцируется одним короткодневным фотопериодом, для фотоиндукции зацветания достаточно нескольких квадратных сантиметров листа. Основную роль в во 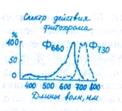 сприятии фотопериода листом играет фитохром. Предполагается, что его темновое превращение (см. рис. 2.15) может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма.
сприятии фотопериода листом играет фитохром. Предполагается, что его темновое превращение (см. рис. 2.15) может служить способом измерения времени по типу песочных часов. Вспышка красного света устанавливает фазу эндогенного ритма.
Необходимость большой поверхности листьев и достаточной интенсивности света для перехода к цветению у многих растений объясняется потребностью растущих меристем в ассими-лятах. Устранение СО2 из воздуха во время светового периода снижает инициацию цветения у некоторых КДР и ДДР.
У фотопериодически нейтральных растений переход к зацветанию обеспечивается возрастными изменениями (эндогенная регуляция).
Эвокация (от лат. evocatio — вызывание) представляет собой завершающую фазу инициации цветения, во время которой в апексе происходят процессы, необходимые для инициации цветочных зачатков. При поступлении флорального стимула в апикальных меристемах последовательно возрастают содержание растворимых Сахаров и активность инвертазы, число митохондрий и интенсивность дыхания; активируется митоз как в периферической, так и центральной зонах стеблевой меристемы; наблюдается кратковременная синхронизация клеточных делений; усиливается синтез РНК и белков, изменяется их качественный состав (последнее — за сутки до заложения зачатков цветков). Все эти данные показывают, что сущность эвокации заключается в переключении генетической программы, способствующей развитию вегетативных почек, на другую, обеспечивающую закладку и формирование цветков.
Природа флорального стимула. Немецкий ботаник Г. Клебс (1903) выдвинул теорию, согласно которой половое размножение всех групп растений (от водорослей до высших) зависит от условий питания. В его опытах такие водоросли, как Сlату-domonas, Vaucheria, и даже некоторые высшие растения (Glec-homa hederata), не проявляли никаких признаков перехода к половому размножению, если постоянно поддерживались благоприятные для них условия существования: состав питательной среды, освещение, температура. Изменение же этих условий в неблагоприятную для вегетативного роста сторону способствовало переходу к половому размножению.
В 1913 г. Клебс высказал предположение, что цветению высших растений способствует высокое соотношение содержания эндогенных углеводов и азотистых соединений (C/N). Действительно, неблагоприятные факторы среды часто ускоряют цветение, а высокий уровень питания, особенно азотного, у некоторых растений вызывает усиление вегетативного роста и задержку генеративного развития. Однако последующее открытие фотопериодизма, выяснение роли фитохрома и фитогормонов в процессах роста и развития показали, что регуляция перехода растений к цветению — гораздо более сложное явление.
 Было установлено, что фотопериодическое воздействие воспринимается листьями и затем флоральный стимул передается в апекс побега. Эти факты позволили М. X. Чайлахяну (1937) сформулировать гормональную теорию развития растений. Согласно этой теории при благоприятном фотопериоде в листьях образуется гормон цветения флориген, индуцирующий переход растений к зацветанию. Эта точка зрения подтверждалась опытами с прививками: зацветание вегетирующих растений, привитых на дветущие, происходило при неблагоприятном фотопериоде за счет веществ, передвигающихся из листьев подвоев. Химически флориген не является видоспеци-фичным, так как цветущие растения одного вида способны индуцировать цветение у привитых вегетирующих растений другого вида.
Было установлено, что фотопериодическое воздействие воспринимается листьями и затем флоральный стимул передается в апекс побега. Эти факты позволили М. X. Чайлахяну (1937) сформулировать гормональную теорию развития растений. Согласно этой теории при благоприятном фотопериоде в листьях образуется гормон цветения флориген, индуцирующий переход растений к зацветанию. Эта точка зрения подтверждалась опытами с прививками: зацветание вегетирующих растений, привитых на дветущие, происходило при неблагоприятном фотопериоде за счет веществ, передвигающихся из листьев подвоев. Химически флориген не является видоспеци-фичным, так как цветущие растения одного вида способны индуцировать цветение у привитых вегетирующих растений другого вида.
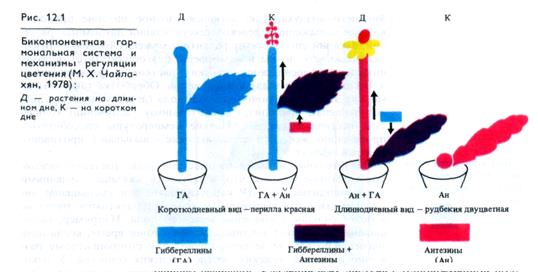
А. Ланг (США) в 1956 г. установил, что цветение многих ДДР значительно ускоряется благодаря обработке гибберелли-ном. Однако в дальнейшем оказалось, что под действием FAj активируется прежде всего развитие цветоносов. Чайлахян (1958) выдвинул гипотезу о бикомпонентной природе флориге-на, согласно которой гормон цветения состоит из гиббереллинов и антезинов, причем гиббереллины обусловливают образование и рост цветочных стеблей, а антезины индуцируют заложение цветков. По этой гипотезе ДДР, особенно розеточные формы, содержат антезины, но бедны гиббереллинами. В условиях благоприятного длиннодневного фотопериода в листьях ДДР синтезируется значительное количество гиббереллинов, которые поступают в апекс побега. Присутствие обоих компонентов флоригена — гиббереллина и антезина — индуцирует зацветание растений (рис. 12.1). У КДР, которые, как правило, имеют высокий стебель, наоборот, гиббереллинов достаточно при любом фотопериоде, а антезин синтезируется только в условиях короткодневного фотопериода. Этим объясняется благоприятное действие короткодневного фотопериода на зацветание КДР. У НДР содержание гиббереллинов и антезинов увеличивается с возрастом до какого-то критического уровня, который необходим для инициации цветения. По бикомпонентной гипотезе флоригена предполагается участие и других известных в настоящее время фитогормонов в регуляции закладки и формирования цветков у всех фотопериодических групп. Теория флоригена в современном ее виде достаточно убедительно объясняет механизм регуляции перехода в генеративное состояние. Однако природа антезина остается неизвестной.
Ж. Бернье, Ж.-М. Кине и Р. Сакс (1985) выдвинули свою гипотезу индукции и эвокации цветения. По мнению этих авторов, эвокация контролируется не одним каким-то специфическим морфогеном, а сложной системой из нескольких факторов, каждый из которых запускает свою цепь эвокационных процессов. Взаимодействие этих процессов и приводит к закладке цветков. Факторы, участвующие в регуляции эвокации, образуются в разных частях растительного организма, причем они не обязательно идентичны у всех видов.
К сказанному нужно добавить, что в листьях растений в условиях неблагоприятного фотопериода обнаруживаются ингибиторы цветения.
 Удаление всех листьев с одновременной подкормкой сахарозой способствует цветению некоторых ДДР и КДР в строго неиндуктивных условиях. При благоприятном фотопериоде концентрация этих ингибиторов снижается, что и приводит к зацветанию растений.
Удаление всех листьев с одновременной подкормкой сахарозой способствует цветению некоторых ДДР и КДР в строго неиндуктивных условиях. При благоприятном фотопериоде концентрация этих ингибиторов снижается, что и приводит к зацветанию растений.
Под детерминацией (определением) пола у растений понимают формирование клеток, органов или особей определенного пола в зависимости от генетических факторов, локализованных в хромосомах (генетическое определение пола), и от условий внешней и внутренней среды (фенотипическое определение пола).
Цветки как органы полового размножения могут быть обоеполыми или реже раздельнополыми. Они формируются на одних и тех же (однодомность) или на разных (двудомность) растениях.
Двудомных видов мало. К ним относятся конопля, хмель, тополь, ива. У некоторых двудомных видов в клетках обнаружены половые хромосомы типа XX и XY (некоторые мхи, элодея, дрема). Генетической основой определения пола у этих двудомных растений являются Х- и Y-хромосомы, однако гены, ответственные за детерминацию пола, локализуются не только в половых хромосомах, но и в аутосомах (т. е. во всех других хромосомах). Тем более это относится к растениям, у которых специализированных половых хромосом нет. Гены, ответственные за определение пола, играют важную роль, позволяя клеткам no-разному реагировать на действие факторов внутренней и внешней среды. В отличие от животных процесс формирования пола в ходе онтогенеза растений может сильно меняться в зависимости от условий вплоть до превращения одного пола в другой.
Проявление пола у растений зависит от таких факторов внешней среды, как длина дня, интенсивность и спектральный состав света, температура, минеральное питание, состав окру  жающего воздуха. Так, хорошее азотное питание приводит к более выраженной женской сексуализации двудомных растений, а калий способствует развитию мужских растений. Высокая влажность почвы и атмосферы благоприятствует заложению и дифференцировке женских цветков и формированию женских растений у двудомных видов. Обработка такими газами, как этилен или монооксид углерода (действующий как аналог этилена), приводит к значительному увеличению количества пестичных цветков. Низкие температуры способствуют проявлению женского пола, высокие — вызывают противоположный эффект.
жающего воздуха. Так, хорошее азотное питание приводит к более выраженной женской сексуализации двудомных растений, а калий способствует развитию мужских растений. Высокая влажность почвы и атмосферы благоприятствует заложению и дифференцировке женских цветков и формированию женских растений у двудомных видов. Обработка такими газами, как этилен или монооксид углерода (действующий как аналог этилена), приводит к значительному увеличению количества пестичных цветков. Низкие температуры способствуют проявлению женского пола, высокие — вызывают противоположный эффект.
Значительное влияние на сексуализацию растений оказывают световые условия, что, в частности, связано с явлениями фотопериодизма. У КДР короткий день или уменьшение интенсивности света вызывает быстрое репродуктивное развитие и более сильное проявление женского пола. Например, выращивание кукурузы в теплице в осенне-зимнее время, когда день значительно короче летнего, приводит к стопроцентному развитию на месте мужских метелок женских соцветий. У некоторых растений в этих условиях усиливается женская сексуали-зация, у других — мужская. Характер дифференциации пола зависит не только от фотопериода, но и от спектрального состава света. Коротковолновый свет усиливает проявление женских признаков, а длинноволновый тормозит этот процесс.
Регуляция пола у раздельнополых растений контролируется фитохромной системой и связана с содержанием и активностью фитогормонов. Действие факторов внешней среды в значительной степени осуществляется через изменение гормонального обмена. Исследования влияния фитогормонов и ретардантов на сексуализацию дают основание для достаточно определенных выводов о роли гиббереллинов как гормонов, усиливающих маскулинизацию (мужскую сексуализацию), и о роли цитокининов, а также ауксина и этилена как фитогормонов, усиливающих феминизацию (женскую сексуализацию) растений.
В связи с изучением роли фитогормонов в проявлении пола следует подробнее остановиться на значении в этом процессе синтезирующих их органов. М. X. Чайлахян с сотрудниками (1977— 1982) показали, что как у двудомных, так и однодомных растений удаление корней, синтезирующих цитокинины, способствует усилению мужской сексуализации (рис. 12.2). Удаление листьев приводит к противоположному эффекту, а одновременное введение гиббереллинов вновь увеличивает число мужских растений. Таким образом, корни влияют на сексуализацию растений как органы, синтезирующие цитокинины, а листья — как органы, синтезирующие гиббереллины. В связи с этим становится понятным, почему мужские экземпляры тополей при ежегодной подрезке крон (уменьшении массы листьев) превращаются в деревья с женской сексуализацией, образующие к досаде горожан большое количество тополиного пуха.
Механизм действия фитогормонов на сексуализацию связан с изменением активности генного аппарата, о чем свидетельствуют опыты по изучению влияния ингибиторов синтеза нуклеиновых кислот и белка на проявление пола растений.

 Таким образом, процесс дифференциации пола можно представить следующим образом. Все факторы внешней среды так или иначе влияют на гормональный обмен растений, причем факторы, приводящие к сдвигу гормонального баланса в сто рону увеличения содержания цитокинина (или в ряде случаев ауксина) усиливают женскую сексуализацию, а вызывающие увеличение содержания гиббереллина — мужскую. Гормональ ная система взаимодействует с генетическим аппаратом, что и обусловливает проявление пола у растений. 1 В ходе эвокации под действием флорального стимула в апи кальной меристеме стебля индуцируется экспрессия генов, от ветственных за морфогенетическую программу цветообразова- ния. Увеличивается число клеточных делений, изменяется форма апекса. Первыми закладываются чашелистики, по являются примордии лепестков, затем последовательно закладываются тычинки и плодолистики. Прежде всего, как и при формировании примордиев листа, физиологически и морфологически выделяются группы инициальных клеток, особенности деления которых определяют дальнейший ход морфогенеза каждой части цветка. Например, тычиночная нить образуется за счет апикального, а затем интеркалярного деления и роста клеток примордия. В дистальной части инициальные клетки дают начало тканям пыльника.
Таким образом, процесс дифференциации пола можно представить следующим образом. Все факторы внешней среды так или иначе влияют на гормональный обмен растений, причем факторы, приводящие к сдвигу гормонального баланса в сто рону увеличения содержания цитокинина (или в ряде случаев ауксина) усиливают женскую сексуализацию, а вызывающие увеличение содержания гиббереллина — мужскую. Гормональ ная система взаимодействует с генетическим аппаратом, что и обусловливает проявление пола у растений. 1 В ходе эвокации под действием флорального стимула в апи кальной меристеме стебля индуцируется экспрессия генов, от ветственных за морфогенетическую программу цветообразова- ния. Увеличивается число клеточных делений, изменяется форма апекса. Первыми закладываются чашелистики, по являются примордии лепестков, затем последовательно закладываются тычинки и плодолистики. Прежде всего, как и при формировании примордиев листа, физиологически и морфологически выделяются группы инициальных клеток, особенности деления которых определяют дальнейший ход морфогенеза каждой части цветка. Например, тычиночная нить образуется за счет апикального, а затем интеркалярного деления и роста клеток примордия. В дистальной части инициальные клетки дают начало тканям пыльника.
 Примордии плодолистиков приобретают подковообразную форму, удлиняются и срастаются краями. Таким образом образуется пестик. На внутренней стороне каждого плодолистика закладывается примордии семяпочки (см. рис. 11.1). Семяпочка разрастается, формируя интегументы и нуцеллус (см. рис. 11.2). Нуцеллус выполняет функции мегаспорангия. Одна из его клеток становится археспориальной. Она делится редукционным путем (мейоз), в результате чего образуются четыре гаплоидные макроспоры. Три из них разрушаются, а одна дает начало зародышевому мешку (женскому гаметофиту). В результате трех последовательных митотических делений в зародышевом мешке появляются 8 гаплоидных ядер, причем после первого деления два дочерних ядра перемещаются к противоположным полюсам поляризованного гаметофита и там делятся еще два раза (см. рис. 11.1). Три ядра у каждого полюса обособляются в клетки. На микропилярном полюсе одна из клеток становится яйцеклеткой (женской гаметой), а две другие — синергидами. Три ядра на халазальном полюсе образуют клетки, называемые антиподами. Два оставшихся ядра мигрируют к центру и сливаются, превращаясь во вторичное диплоидное ядро центральной клетки, которая дает начало эндосперму (см. рис. 11.1 и 11.2). В таком виде яйцеклетка и женский гаметофит в целом готовы для оплодотворения.
Примордии плодолистиков приобретают подковообразную форму, удлиняются и срастаются краями. Таким образом образуется пестик. На внутренней стороне каждого плодолистика закладывается примордии семяпочки (см. рис. 11.1). Семяпочка разрастается, формируя интегументы и нуцеллус (см. рис. 11.2). Нуцеллус выполняет функции мегаспорангия. Одна из его клеток становится археспориальной. Она делится редукционным путем (мейоз), в результате чего образуются четыре гаплоидные макроспоры. Три из них разрушаются, а одна дает начало зародышевому мешку (женскому гаметофиту). В результате трех последовательных митотических делений в зародышевом мешке появляются 8 гаплоидных ядер, причем после первого деления два дочерних ядра перемещаются к противоположным полюсам поляризованного гаметофита и там делятся еще два раза (см. рис. 11.1). Три ядра у каждого полюса обособляются в клетки. На микропилярном полюсе одна из клеток становится яйцеклеткой (женской гаметой), а две другие — синергидами. Три ядра на халазальном полюсе образуют клетки, называемые антиподами. Два оставшихся ядра мигрируют к центру и сливаются, превращаясь во вторичное диплоидное ядро центральной клетки, которая дает начало эндосперму (см. рис. 11.1 и 11.2). В таком виде яйцеклетка и женский гаметофит в целом готовы для оплодотворения.
В пыльнике клетки спорогенной ткани последовательно делятся два раза — путем мейоза, а затем митоза; в результате из каждой материнской клетки образуются четыре гаплоидные микроспоры (см. рис. 11.1). В период образования микроспор спорогенные материнские клетки теряют большую часть ци-топлазматических РНК и белков. При прорастании ядро микроспоры делится митотически и асимметрично, в результате чего внутри пыльцевого зерна (мужского гаметофита) с вегетативным ядром появляется маленькая генеративная клетка со своим генеративным ядром.
Основная роль синергид заключается в привлечении пыльцевой трубки к зародышевому мешку путем выделения хемо-тропных веществ и в проведении спермиев в пространство между яйцеклеткой и центральной клеткой. Возможно также, что синергиды выполняют функции гаусторий, поглощая питательные вещества из соматических клеток и снабжая ими яйцеклетку и зародышевый мешок. Роль антипод состоит в обеспечении зародышевого мешка питательными веществами.
Центральный момент репродуктивного развития — индукция мейоза. Успешное завершение мейоза в культивируемых изолированных пыльниках в условиях эксперимента возможно, если споровые материнские клетки во время изоляции пыльника уже вступили в мейоз. В этом случае благотворный эффект на образование тетрад и последующие стадии развития одноядерных микроспор оказывают среды с сахарами, минеральными солями и цитокинином, гиббереллином, РНК или ее нуклеотидами.
Если пыльники отрезать до начала или во время митоза пыльцевых зерен и культивировать в простой минеральной среде с добавлением сахара, то большая часть микроспор становится нежизнеспособной, у другой части вместо нормального гаметогенеза начинаются клеточные деления с образованием гаплоидных проростков (пыльцевых эмбриоидов).

 В процессе мега- и микроспорогенеза в клеточных стенках на ранних фазах развития откладывается каллоза (β-1,3-глю-кан), обеспечивающая временную изоляцию клеток, переходящих на генеративный путь развития. Генеративная клетка при формировании также на какое-то время покрывается клеточной оболочкой из каллозы, что необходимо для ее изоляции от цитоплазмы вегетативной клетки. Затем каллоза исчезает и генеративная клетка остается окруженной оболочкой из целлюлозы и пектина. Таким образом, временная изоляция клеток — важное условие для перехода к генеративной фазе развития.
В процессе мега- и микроспорогенеза в клеточных стенках на ранних фазах развития откладывается каллоза (β-1,3-глю-кан), обеспечивающая временную изоляцию клеток, переходящих на генеративный путь развития. Генеративная клетка при формировании также на какое-то время покрывается клеточной оболочкой из каллозы, что необходимо для ее изоляции от цитоплазмы вегетативной клетки. Затем каллоза исчезает и генеративная клетка остается окруженной оболочкой из целлюлозы и пектина. Таким образом, временная изоляция клеток — важное условие для перехода к генеративной фазе развития.
Генеративная клетка делится и образует два спермия (мужские гаметы). У многих видов это происходит уже в пыльцевой трубке. Вегетативная клетка обеспечивает жизнедеятельность пыльцевого зерна и рост пыльцевой трубки.
Оплодотворение — один из важнейших процессов полового размножения. Его можно подразделить на три фазы: опыление, прорастание пыльцы и рост пыльцевой трубки в тканях пестика и собственно оплодотворение, т. е. образование зиготы. Анатомо-морфологически эти процессы исследованы у многих покрытосеменных. Физиолого-биохимические аспекты оплодотворения в настоящее время интенсивно изучаются.
Для того чтобы произошло опыление, пыльца с помощью насекомых, ветра или воды должна попасть на рыльце пестика цветка. Различают пыльцевые зерна с клейкой и неклейкой поверхностью. Оболочка пыльцевого зерна состоит из внутреннего и наружного слоев: интины и экзины соответственно. Ин-тина, помимо целлюлозы и пектиновых веществ, содержит белки, участвующие в процессах взаимного «узнавания» пыльцы и тканей рыльца, и гидролитические ферменты (кислую фосфатазу, протеазу, РНКазу и др.). Они концентрируются вблизи поры или пор прорастания и активируются при контакте поры с тканью рыльца. Экзина состоит из спорополлени-на — вещества терпеноидной природы и обладает высокой устойчивостью к внешним воздействиям. В полостях экзины локализованы белки, которые функционируют в системе несовместимости пыльцы и рыльца, контролируя внутривидовую и межвидовую совместимость при опылении. Пыльцевое зерно содержит ряд запасных веществ (крахмал, липиды, белки), витамины, а также активаторы и ингибиторы роста. Зрелая пыльца содержит лишь 10—15% воды.
При попадании на рыльце пестика пыльца начинает набухать. Рыльца зрелых пестиков могут быть покрыты секретом или не содержат его («сухие рыльца»). Железистая ткань рыльца, выделяющая секрет, состоит из эпидермальных клеток, вытянутых в форме папилл (сосочков). Секрет вырабатывается и в субэпидермальных слоях клеток рыльца. Он состоит из липидов (производных восков), выполняющих функцию защиты от потери воды, и фенольных соединений (антоцианов, флаво-ноидов, коричных кислот). Фенольные компоненты могут участвовать в регуляции прорастания пыльцы, в защите от инфекций и в системе распознавания совместимости. У ряда растений секрет содержит также разнообразные белки.
На рыльце одновременно может попадать много пыльцевых зерен, но прорастают лишь совместимые. Механизм несовместимости до конца не выяснен, но известно, что при контакте пыльцевого зерна с рыльцем и его набухании из экзины и интины начинают выделяться содержащиеся в них белки (гликопротеины). При контакте белков экзины с несовместимым рыльцем сразу начинается образование каллозы, изолирующей пыльцевое зерно. Прорастание несовместимой пыльцы может остановиться не только на поверхности, но и в ткани рыльца или в столбике. Так, рыльце секретирует S-факторы (гликопротеины), которые, взаимодействуя с компонентами клеточной стенки несовместимой пыльцевой трубки, подавляют ее рост.
В случае совместимого рыльца активируется кутиназа и другие гидролитические ферменты интины, разрыхляющие стенки клеток рыльца и способствующие росту пыльцевой трубки. При этом начальные этапы проникновения пыльцевой трубки определяются ее положительной гидротропической реакцией, а на конечных стадиях рост пыльцевой трубки представляет собой положительную хемотропическую реакцию на вещества, выделяемые тканями микропиле, синергидами и яйцеклеткой. Сведения о веществах, необходимых для роста пыльцевых трубок, были получены при проращивании пыльцы в условиях in vitro. Пыльца большинства видов хорошо прорастает в присутствии сахарозы, различных минеральных веществ, в том числе ионов кальция; в ряде случаев требуется добавление аминокислот и гиббереллина. По-видимому, все эти соединения пыльцевая трубка получает из рыльца и из тканей столбика.
Растущая пыльцевая трубка перемещается в столбике между клетками или — у некоторых видов — по пектиновым слоям в толстых клеточных стенках. Проводниковая ткань столбика способствует росту пыльцевой трубки, снабжая ее питательными веществами и активаторами роста.
Пыльцевые трубки секретируют ферменты, разрыхляющие кутин рыльца и пектиновые вещества срединных пластинок клеточных стенок, что позволяет пыльцевой трубке перемещаться в столбике. Кроме того, пыльцевые трубки выделяют ауксин (или фактор, ускоряющий его синтез), участвующий в инициации развития плода. Однако резкое возрастание уровня ауксина в тканях пестика наблюдается сразу после опыления, что не может быть связано с привнесением биологически активных веществ только с пыльцой.
Непосредственно после опыления наблюдается приток фосфора к завязи, изменение распределения в ней белков и углеводов. Все эти явления нельзя объяснить непосредственным воздействием пыльцевых трубок. Возможно, в регуляции подобных процессов большую роль играют электрофизиологические процессы. При опылении цветков в рыльцах появляется потенциал действия, распространяющийся в направлении завязей. ПД, по-видимому, выполняет сигнальную функцию, так как завязь отвечает на электрофизиологический сигнал изменением метаболизма, с которого начинается подготовка женской репродуктивной системы к оплодотворению.
 Пыльцевая трубка, пройдя через микропиле, через одну из синергид проникает в зародышевый мешок; спермин выходят и один из них сливается с ядром яйцеклетки, образуя зиготу, а другой — с полярными ядрами (или вторичным ядром) центральной клетки, дающей начало эндосперму (двойное оплодотворение, открытое в 1898 г. С. Г. Навашиным). Значение двойного оплодотворения состоит в том, что как зигота, так и первичная клетка эндосперма, имея двойную наследственность, приобретают большую жизнеспособность и приспособляемость к условиям существования. Отсюда понятна роль многочисленных приспособлений в морфологии и физиологии цветка, направленных на обеспечение перекрестного опыления. В зародышевый мешок могут проникать несколько пыльцевых трубок. Однако спермин этих трубок в оплодотворении, как правило, не участвуют и дегенерируют. Слиянию спермиев с ядрами женского гаметофита предшествует деконденсация хроматина в ядрах.
Пыльцевая трубка, пройдя через микропиле, через одну из синергид проникает в зародышевый мешок; спермин выходят и один из них сливается с ядром яйцеклетки, образуя зиготу, а другой — с полярными ядрами (или вторичным ядром) центральной клетки, дающей начало эндосперму (двойное оплодотворение, открытое в 1898 г. С. Г. Навашиным). Значение двойного оплодотворения состоит в том, что как зигота, так и первичная клетка эндосперма, имея двойную наследственность, приобретают большую жизнеспособность и приспособляемость к условиям существования. Отсюда понятна роль многочисленных приспособлений в морфологии и физиологии цветка, направленных на обеспечение перекрестного опыления. В зародышевый мешок могут проникать несколько пыльцевых трубок. Однако спермин этих трубок в оплодотворении, как правило, не участвуют и дегенерируют. Слиянию спермиев с ядрами женского гаметофита предшествует деконденсация хроматина в ядрах.
Плод развивается из цветка и, как правило, содержит семена. Образование семян из семяпочки уже обсуждалось в разделе 11.1. Ткани плода формируются из завязи, но нередко в образовании плода участвуют и другие части цветка, например цветоложе, околоцветник, цветковые чешуи. Ткани плода закладываются при формировании цветка, причем в ряде случаев плод может развиваться и независимо от семян (партенокарпи-ческие, т. е. бессемянные, плоды). Однако обычно цветки без опыления и оплодотворения опадают.
Развитие плодов можно разделить на четыре фазы: 1) формирование завязи до опыления; 2) рост за счет деления клеток сразу после опыления и оплодотворения; 3) рост за счет растяжения клеток; 4) созревание.
Резкое усиление деления клеток завязи наблюдается вскоре после опыления. Затем наступает фаза растяжения клеток. Характер роста находится в сильной зависимости от типа плода. Сигмоидная кривая роста плодов характерна, например, для томатов, яблок, авокадо и др. В этих случаях клетки делятся почти исключительно во время формирования цветочной почки. После опыления деление клеток еще некоторое время продолжается (этот период занимает от нескольких дней у томатов до 1,5 месяцев у некоторых сортов груш). Дальше увеличение размеров плода идет за счет растяжения клеток.
У плодов типа костянок (абрикос, слива, вишня и др.) рост характеризуется двойной сигмоидной кривой. Первый быстрый период роста обусловлен разрастанием завязи, нуцеллуса и интегументов семени, а зародыш и эндосперм в это время практически не развиваются. Когда же во второй фазе начинается развитие эндосперма и зародыша, завязь растет очень слабо. В это же время происходит склерификация косточки. Когда зародыш достигает полной зрелости, начинается второе ускорение роста плода, которое продолжается до полного его созре вания. Таким образом, наблюдается взаимозависимость между развитием семян и ростом плодов.


Деление клеток будущего плода зависит от веществ, вырабатываемых листьями в период, предшествующий цветению. Размеры меристем, обусловливающие размер плодов, сильно зависят от снабжения их углеводами и азотом, т. е. от метаболизма целого растения.
Регуляция следующего этапа развития плодов связана с опылением. Одно только помещение даже чужеродной пыльцы на рыльце, без последующего оплодотворения, индуцирует разрастание стенки завязи без развития семян. Это действие можно заменить и мертвой пыльцой, и экстрактами из нее. Факторы, индуцирующие разрастание завязи и содержащиеся в экстрактах пыльцы, не являются видоспецифичными и представляют собой вещества гормональной природы. Это ауксины и гиббереллиноподобные соединения. Кроме того, при опылении повышается способность тканей пестика синтезировать собственный ауксин.
Оплодотворенные яйцеклетка и эндосперм и уже начавшие развиваться семена оказывают сильное контролирующее влияние на рост плодов. Так, недоразвитие семян в силу каких-либо причин (например, воздействие паразитических организмов) служит причиной преждевременного опадения плодов. Неравномерное развитие семян в многосемянных плодах вызывает изменение их формы, развитие асимметричных, неправильной формы плодов (рис. 12.3).
То, что семяпочки и развивающиеся семена регулируют рост плодов посредством вырабатываемых ими гормонов, подтверждается многими данными. Для различных растений показано, что развивающиеся семена являются центрами синтеза ауксина, гиббереллина и цитокининов. Именно из незрелых семян впервые были выделены ГА8 и зеатин. Эти регулирующие рост вещества делают развивающийся плод центром притяжения (акцептором) питательных веществ, синтезируемых в листьях, и развитие плодов обычно связано с заметным прекращением вегетативного роста, а у однолетников — со старением всего растения. Удаление плодов задерживает старение листьев. Так, у фасоли перемещение 32Р и углерода 14С из листьев к плодоножке вызывается развитием стручков или обработкой плодоножки с удаленным стручком ланолиновой пастой с ИУК, ГА и кинетином. При недостатке ауксина плоды опадают.
 Концепция о том, что регулирующие рост вещества, сущеественные для роста плода (ауксин, гиббереллины, цитокинин), синтезируются в оплодотворенных семяпочках, поддерживается опытами с культурой изолированных цветков. Развитие из изолированного цветка плода до его созревания осуществляется у ряда видов на простой среде из минеральных веществ и сахарозы, если цветки были взяты спустя два дня и более после опыления материнского растения. Из цветков неопыленных томатов получали партенокарпические плоды при добавлении в культуральную среду 2-нафтоксиуксусной кислоты. Если в среду вносили томатный сок (содержащий цитокинин), то вырастали крупные плоды.
Концепция о том, что регулирующие рост вещества, сущеественные для роста плода (ауксин, гиббереллины, цитокинин), синтезируются в оплодотворенных семяпочках, поддерживается опытами с культурой изолированных цветков. Развитие из изолированного цветка плода до его созревания осуществляется у ряда видов на простой среде из минеральных веществ и сахарозы, если цветки были взяты спустя два дня и более после опыления материнского растения. Из цветков неопыленных томатов получали партенокарпические плоды при добавлении в культуральную среду 2-нафтоксиуксусной кислоты. Если в среду вносили томатный сок (содержащий цитокинин), то вырастали крупные плоды.
Кроме перечисленных выше гормонов в плодах и семенах обнаруживаются и другие биологически активные вещества, регулирующие их развитие. Так, в плодах найдены регуляторы клеточных делений — цитокинины, а также ингибиторы роста, такие, как салициловая, я-кумаровая, феруловая и другие кислоты. Присутствуют также ингибиторы прорастания семян, такие, как абсцизовая кислота. Уровень и соотношение гормонов и ингибиторов от времени оплодотворения до созревания плодов сильно изменяется. Так, вскоре после оплодотворения (через неделю в завязи фасоли, например) наблюдается максимум активности гиббереллинов. Немного позже максимальной активности достигают ауксины. Увеличение содержания ауксина совпадает с переходом эндосперма к клеточному делению. С момента достижения максимума концентрации ауксина начинает активно расти зародыш. Следующий пик активности ауксина связан с усилением клеточных делений на периферии эндосперма, где и содержится максимальное количество ауксина в уже сформировавшихся семенах. Наивысший уровень регуляторов клеточных делений характерен для очень молодых плодов и семян.
Созревание плодов сопровождается усиленным продуцированием этилена, который резко ускоряет эту завершающую фазу их развития.
При развитии плодов и семян происходят значительные метаболические изменения. Так, дыхание плодов, очень интенсивное на первых этапах развития, снижается по мере их роста и кратковременно усиливается у многих сочных плодов в период созревания (климактерический период). Основными метаболитами, потребляемыми плодами в процессах дыхания, являются сахара и органические кислоты. Их содержание в ходе развития плодов сильно меняется. Уже через несколько дней после опыления или обработки ауксином в плодах резко снижается концентрация Сахаров. Во время фаз быстрого развития плодов и семян идет интенсивный синтез крахмала, гидролизуемого в период созревания. В плодах некоторых растений крахмал вовсе отсутствует. В разных плодах обнаруживаются также глюкуроновая кислота, сорбитол и др. Углеводы частично синтезируются в хлоропластах, которые присутствуют в незрелых плодах, однако в основном ассимиляты поступают в плоды из фотосинтезирующих листьев.
Среди органических кислот в плодах преобладают ди и трикарбоновые, такие, как лимонная, яблочная, винная и др. В большом количестве в молодых плодах обнаруживаются ароматические кислоты. Во время роста плодов содержание органических кислот увеличивается, а при их созревании опять уменьшается.
 Показано, что накопление органических кислот зависит от климатических факторов: при солнечной погоде органические кислоты используются на дыхание, при пасмурной — накапливаются. Вопрос об образовании органических кислот в плодах до конца не ясен. По некоторым данным, уменьшение листовой поверхности ведет к снижению содержания кислот в плодах, что может свидетельствовать о синтезе их в листьях с последующим перемещением в плоды. По другим данным, незрелые плоды способны к темновой фиксации СO2, который включается в яблочную, лимонную и другие кислоты.
Показано, что накопление органических кислот зависит от климатических факторов: при солнечной погоде органические кислоты используются на дыхание, при пасмурной — накапливаются. Вопрос об образовании органических кислот в плодах до конца не ясен. По некоторым данным, уменьшение листовой поверхности ведет к снижению содержания кислот в плодах, что может свидетельствовать о синтезе их в листьях с последующим перемещением в плоды. По другим данным, незрелые плоды способны к темновой фиксации СO2, который включается в яблочную, лимонную и другие кислоты.
Процессы созревания начинаются, когда плоды заканчивают свой рост. Созревание сочных плодов характеризуется снижением соотношения кислот и Сахаров, образованием ароматических веществ, разложением хлорофилла и дубильных веществ, накоплением антоцианов и других вакуолярных пигментов, уменьшением твердости и упругости тканей благодаря гидролизу пектиновых веществ клеточных стенок. Однако на начальных этапах созревания в плодах еще преобладают процессы синтеза веществ. После прохождения периода созревания плод вступает в фазу отмирания.
 Вегетативное размножение — типичное свойство растений, отличающее их от животных. Это — воспроизведение растений из их вегетативных частей. Наиболее сложных и разнообразных форм оно достигает у высших и особенно у цветковых растений. Существуют три основных способа вегетативного размножения, используемых в естественных условиях: 1) размножение отпрысками различной природы, отделяющимися от материнской особи после укоренения, 2) размножение клубнями и луковицами, 3) обособление частей материнской особи.
Вегетативное размножение — типичное свойство растений, отличающее их от животных. Это — воспроизведение растений из их вегетативных частей. Наиболее сложных и разнообразных форм оно достигает у высших и особенно у цветковых растений. Существуют три основных способа вегетативного размножения, используемых в естественных условиях: 1) размножение отпрысками различной природы, отделяющимися от материнской особи после укоренения, 2) размножение клубнями и луковицами, 3) обособление частей материнской особи.
Вегетативное размножение растений имеет огромное биологическое значение. Так, способность покрытосеменных к интенсивному вегетативному размножению сыграла немалую роль в их успешной конкуренции с голосеменными, которым свойственно только семенное размножение. Во многих случаях вегетативное размножение становится единственно возможной формой репродукции, обеспечивающей расселение вида и сохранение генофонда. Многие растения со специализированными органами вегетативного размножения практически утрачивают способность к размножению семенами, передавая функции как размножения, так и запасания питательных веществ этим органам.
Между развитием специализированных органов вегетативного и полового размножения наблюдается ряд аналогий. И те и другие органы образуются в результате индукции, воспринимаемой листьями и передающейся при помощи химических стимулов соответствующим частям растения. После дифференцировки и роста в обоих случаях наступает этап созревания. Морфологически как цветки, так и клубни и луковицы являяются видоизменениями побега. Клубень представляет собой утолщенную вздутую часть стебля, состоящую из нескольких междоузлий. Луковицы образуются вследствие укорочения междоузлий и иммобилизации углеродов в основаниях очень молодых листьев. При образовании клубней и луковиц, как и при цветении, вегетативный рост подавляется.
Сходство специализированных органов вегетативного размножения с семенами в биологическом отношении отмечал Н. П. Кренке: например, образование выводковых почек на цветочных стеблях вместо или одновременно с цветками у лука, чеснока и др., формирование клубеньков в соцветиях картофеля и др.. Все основные этапы фотопериодической реакции при образовании органов вегетативного размножения также близки к фотопериодическим реакциям цветущих растений.
 Размножение клубнями. Способность к клубнеобразованию формируется в процессе онтогенеза постепенно и проявляется после завершения ювенильного этапа. В процессе клубнеобра-зования вначале развиваются столоны — стебли с измененной геотропической реакцией (1-я фаза), а затем на них формируются клубни (2-я фаза). Наиболее существенное влияние на клубнеобразование оказывают температура и длина дня. При этом у одной группы растений клубни образуются только в условиях короткого дня, а у другой — как при коротком, так и при длинном дне. В опытах с прививками растений, не образующих клубни, на клубненосы (например, томатов на картофель, подсолнечника на топинамбур) было выяснено, что стимул клубнеобразования у фотопериодически чувствительных клубненосов вырабатывается в листьях, что он не обладает ви-доспецифичностью и имеет гормональную природу. Оказалось, что в этом процессе взаимодействуют основные группы фитогормонов: ауксины подавляют клубнеобразование; гиббереллины, индуцирующие рост стеблей, усиливают рост столонов и тем самым в определенном отношении способствуют росту клубней. Наиболее значительное влияние на формирование клубней оказывают цитокинины. В условиях короткого дня в корнях и столонах наблюдается высокое содержание цитоки-нинов, а в листьях — абсцизинов. При длинном дне в листьях выше содержание гиббереллинов, а в стеблевых почках — ауксинов.
Размножение клубнями. Способность к клубнеобразованию формируется в процессе онтогенеза постепенно и проявляется после завершения ювенильного этапа. В процессе клубнеобра-зования вначале развиваются столоны — стебли с измененной геотропической реакцией (1-я фаза), а затем на них формируются клубни (2-я фаза). Наиболее существенное влияние на клубнеобразование оказывают температура и длина дня. При этом у одной группы растений клубни образуются только в условиях короткого дня, а у другой — как при коротком, так и при длинном дне. В опытах с прививками растений, не образующих клубни, на клубненосы (например, томатов на картофель, подсолнечника на топинамбур) было выяснено, что стимул клубнеобразования у фотопериодически чувствительных клубненосов вырабатывается в листьях, что он не обладает ви-доспецифичностью и имеет гормональную природу. Оказалось, что в этом процессе взаимодействуют основные группы фитогормонов: ауксины подавляют клубнеобразование; гиббереллины, индуцирующие рост стеблей, усиливают рост столонов и тем самым в определенном отношении способствуют росту клубней. Наиболее значительное влияние на формирование клубней оказывают цитокинины. В условиях короткого дня в корнях и столонах наблюдается высокое содержание цитоки-нинов, а в листьях — абсцизинов. При длинном дне в листьях выше содержание гиббереллинов, а в стеблевых почках — ауксинов.
Согласно М. X. Чайлахяну (1984), механизм гормональной индукции клубнеобразования у видов, образующих клубни на коротком дне, включает в себя две фазы (рис. 12.4). Вначале из листьев в нижние стеблевые почки поступают гиббереллины и абсцизины. Поскольку при коротком дне соотношение их сдвинуто в сторону преобладания абсцизинов (АБК/ГА), ускоренный рост столонов, вызываемый гиббереллином, тормозится. Во время второй фазы образуются клубни, рост которых регулируется более высокой концентрацией цитокинина в корнях и столонах по сравнению с ауксином (Цк/ИУК).
В условиях длинного дня при неблагоприятном для клуб-необразования соотношении гормонов (в листьях ГА/АБК, в корнях — ИУК/Цк) происходит образование и рост корней и столонов. Таким образом, стимулом клубнеобразования, вырабатываемым в листьях, служит определенное соотношение гиббереллинов и абсцизинов, влияющее на первую фазу клуб-необразования.
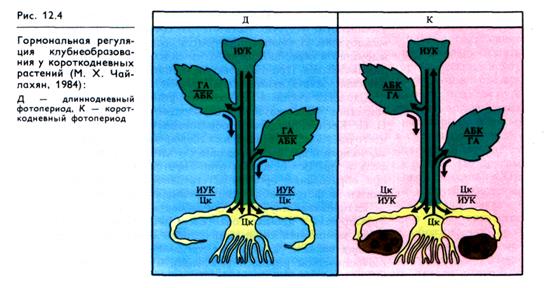
Развитие и рост клубней поддерживается притоком к ним продуктов фотосинтеза, образуемых в листьях. При этом интенсивность транспорта ассимилятов в клубни коррелирует с содержанием в них ауксинов и гиббереллинов, создающих ат-трагирующую силу клубней. Именно поэтому при интенсивном клубнеобразовании замедляется рост надземных частей растения.
У сортов картофеля, выращиваемых в условиях короткого дня, вначале идет рост надземных частей, а затем — интенсивное столоно- и клубнеобразование. Для сортов, образующих клубни и при длинном дне, характерен более протяженный период вегетации, когда развивается надземная масса, а формирование клубней начинается с укорочением дня во второй половине лета.
Размножение луковицами. Если большинство клубненосов образует клубни на коротком дне, то для луковичных растений, напротив, характерно формирование луковиц на длинном дне. Возможно, это связано с эколого-географическим происхождением разных видов. Регуляция образования луковиц изучена гораздо слабее, чем регуляция клубнеобразования. Органами, рецептирующими действие фотопериода, в данном случае также являются листья. В опытах Ф. Э. Реймерса выдерживание даже части листьев на коротком дне замедляло формирование луковицы. Фотопериодический стимул может передаваться от одного побега лука к другому только при условии, что второй побег обезлиствен. Если же побеги с листьями подвергаются воздействию разных фотопериодов, то луковицу формирует лишь побег, находившийся на длинном дне. При образовании луковиц фотопериодический стимул, как при клубнеобразовании и цветении, по-видимому, является комплексом фитогормонов. Так, формирование луковицы сопровождается уменьшением активности ауксинов, а повышение их концентрации ведет к росту листъев и луковичных чешуй в длину. Однако вегетативное размножение луковичных растений связано не столько с формированием луковицы, сколько с образованием на ней пазушных побегов — будущих доячечерних луковичек, «деток». Они закладываются в период зимнего покоя луковиц. Обнаружено, что под действием кинетика, ГА? и ИУК, нанесенных на донце луковицы, значительно усиливается образование почек, формирующих дочерние луковички. Тот же эффект наблюдается при повышении температуры зимнего хранения луковиц или при удалении цветоносного побега.
Прорастание клубней и луковиц происходит по истечении периода покоя. У луковичных растений в отличие от семян и почек, ростовые процессы полностью не прекращаются. В период покоя, хотя и очень медленно, формируются новые органы. Выход из состояния покоя ускоряется после обработки луковиц низкой температурой, т. е. после яровизации. Состояние покоя луковиц поддерживается высоким содержанием в них АБК, а при прорастании снижается уровень ингибиторов и повышается содержание ауксинов, цитокининов и гиббереллинов.
 Знание принципов регуляции состояния покоя клубней и луковиц позволяет предотвращать их прорастание в период хранения, например лука и картофеля, с помощью многих синтетических ингибиторов роста, таких, как гидразид малеиновой кислоты.
Знание принципов регуляции состояния покоя клубней и луковиц позволяет предотвращать их прорастание в период хранения, например лука и картофеля, с помощью многих синтетических ингибиторов роста, таких, как гидразид малеиновой кислоты.
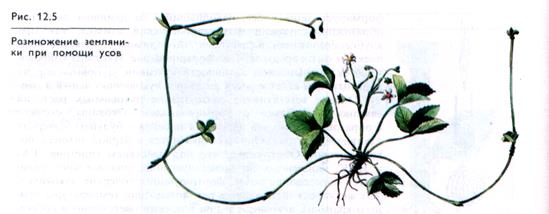
Размножение отводками и другими способами. У дикорастущих растений широко распространено размножение корневой порослью, т. е. отрастанием при повреждении растения спящих почек в нижней части ствола (пневые побеги), корневыми побегами, развивающимися из адвентивных почек на поврежденных корнях у поверхности почвы, корневищами, отводками — ветвями, соприкасающимися с почвой и образующими в месте контакта корни; плетями — у видов с ползучим стеблем, образующим придаточные корни в узлах (лютик ползучий, лапчатка); усами, представляющими собой видоизмененные стебли, из каждой почки которых образуется новое растение. Примером может служить земляника (рис. 12.5), у которой после укоренения узлов происходит формирование самостоятельных растений.
Многие способы вегетативного размножения растений, такие, как размножение при помощи черенков, образование выводковых почек и др., тесно связаны с процессами регенерации (см. 11.3). Так, образование выводковых почек, или вивипария, может рассматриваться как случай соматического эмбриогенеза. Показано, что необходимым условием формирования выводковых почек, например у каланхое, является возникновение физиологической разнокачественности между тканями листьев.
 Эта разнокачественность связана с реакцией на длинный день и выражается в увеличении на краях листа содержания хлорофилла, белкового и небелкового азота, ДНК и РНК. Таким образом, образование выводковых почек как специализированных органов вегетативного размножения контролируется фотопериодически.
Эта разнокачественность связана с реакцией на длинный день и выражается в увеличении на краях листа содержания хлорофилла, белкового и небелкового азота, ДНК и РНК. Таким образом, образование выводковых почек как специализированных органов вегетативного размножения контролируется фотопериодически.
Размножение черенками, прививками и отводками — наиболее распространенные приемы вегетативного размножения, применяемые в растениеводстве.
Размножение отводками заключается в пригибании нижних ветвей растения вниз и засыпании части ветки землей. После того как засыпанный участок дает корни, он отделяется и формируется целое растение. Этот вид размножения близок к размножению усами и плетями, однако укореняющиеся побеги морфологически не отличаются от прочих.
В плодоводстве чаще всего применяют размножение черенками, особенно стеблевыми. Благодаря вегетативному размножению число сортов плодовых деревьев очень велико, причем разница между сортами часто даже больше видовых различий. При размножении черенками следует учитывать, что черенки, взятые из разных участков растения, обладают разными свойствами. Так, черенки, полученные из плодоносной зоны, дают растения, зацветающие и начинающие плодоносить значительно раньше, чем растения, выращенные из ювенильных черенков. Черенки, взятые с бокового побега, иногда долго сохраняют двустороннее строение, не образуя вертикального симметрично построенного растения.
Способность стеблевых черенков к укоренению зависит от многих факторов. В целом черенки от старых деревьев укореняются хуже, чем от молодых. Большое значение может иметь и физиологическое состояние самого черенка. Показано, что максимальной способностью к укоренению обладают побеги в период замедления их вегетативного роста, когда содержание в них эндогенных ауксинов наиболее высокое. Со свойством ауксинов индуцировать корнеобразование связаны методы искусственного повышения способности черенков к укоренению, которая может сильно различаться даже в пределах одного вида. Например, некоторые виды тополя или ивы легко размножаются черенками, тогда как черенки других видов тех же родов практически не дают корней. Трудности размножения видов плохо или совсем неукореняющихся можно преодолеть, обрабатывая черенки различными ауксинами (ИУК, НУК, ИМК и др.), стимулирующими корнеобразование, либо прививая их на растения другого вида, способного к размножению черенками. При помощи прививок развитие черенка значительно ускоряется благодаря корневой системе подвоя. При этом привой сохраняет все свои свойства.
Срастание привоя и подвоя происходит следующим образом. Живые клетки вблизи поверхности среза, особенно клетки камбия, коры, сосудистых пучков, начинают интенсивно делиться, заполняя щель между раневыми поверхностями промежуточной тканью. Через некоторое время клетки привоя и подвоя соединяются плазмодесмами, а затем образуются дифференцированные клетки, соединяющие проводящие системы привоя и подвоя.
В последние годы в практике сельского хозяйства и селекционной работы широко используется метод получения растений с нужными свойствами с помощью культуры тканей. Из одной клетки, группы клеток ткани или суспензии клеток в стерильных условиях выращивают целые растения для клонально-го размножения ценных культур, получают безвирусные формы сельскохозяйственных растений (картофель, сахарная свекла, виноград, древесные породы и др.), путем слияния протопластов создаются межвидовые гибриды, развиваются методы культивирования частей гаметофита и репродуктивных органов древесных и др. Этот метод широко применяется для создания банка безвирусных сортов многих культур.
Половое размножение цветковых растений начинается с индукции цветения, которая может определяться возрастом, а может зависеть от яровизации и (или) фотопериода. По отношению к фотопериодическому воздействию растения делятся на нейтральные, длиннодневные, ко-роткодневные, длиннокороткодневные и др. Фотопериодическое воздействие воспринимается листьями и осуществляется при участии фитохрома. Предполагается, что в листьях образуется гормон цветения — флориген, в состав которого входят гиббереллин и антезин. Длиннодневные растения имеют достаточное количество антезина, но им недостает гиббереллина, который синтезируется на длинном дне. Короткодневные растения, наоборот, не испытывают недостатка в гиббереллине, но им необходим антезин, образующийся в условиях короткодневного фотопериода. Флориген поступает в вегетативные апексы и индуцирует их превращение в апексы флоральные (эвокация). Дифференцировка пола у двудольных растений определяется генотипом и гормональным балансом. Дальнейшие этапы полового размножения включают в себя развитие цветка, опыление и оплодотворение, формирование семян и плодов.
Вегетативные способы размножения очень широко распространены у растений. В некоторых случаях для перехода к вегетативному размножению также необходим благоприятный фотопериодический режим, способствующий созданию в растениях индуктивного гормонального баланса.
 Глава 13
Глава 13

