 2015-05-26
2015-05-26 8856
8856«Три основных вопроса составляют проблему передвижения веществ. Это прежде всего вопрос о тканях, по которым происходит передвижение веществ, вопрос о направлениях передвижения... и третий вопрос — о механизме передвижения».
Д. А. Сабинин
 Ближний транспорт — это передвижение ионов, метаболитов и воды между клетками и тканями (в отличие от мембранного транспорта в каждой клетке). Дальний транспорт — передвижение веществ между органами в целом растении.
Ближний транспорт — это передвижение ионов, метаболитов и воды между клетками и тканями (в отличие от мембранного транспорта в каждой клетке). Дальний транспорт — передвижение веществ между органами в целом растении.
Транспорт веществ в растении осуществляется по любым тканям и по проводящим пучкам, специализированным для этой цели. В свою очередь передвижение воды и растворенных веществ по любым тканям может происходить: а) по клеточным стенкам, т. е. по апопласту, б) по цитоплазме клеток, соединенных друг с другом плазмодесмами, т. е. по симпласту, в) возможно, по эндоплазматическому ретикулуму с участием плазмодесм. Передвижение воды и веществ по проводящим пучкам включает в себя транспорт по ксилеме («восходящий ток» — от корней к органам побега) и по флоэме («нисходящий ток» — от листьев к зонам потребления питательных веществ или отложения их в запас). По флоэме транспортируются метаболиты и при мобилизации запасных веществ.
У многоклеточных водорослей метаболиты передвигаются по симпласту. Исключение составляют ламинарии — крупные бурые водоросли, у которых таллом дифференцирован на ткани и ситовидные трубки формируются в центральной части стебля. Симпластное и апопластное проведение воды и веществ вдоль всего растения присуще и большинству мхов. У всех остальных высших наземных растений имеются проводящие пучки, состоящие из трахеид и (или) сосудов ксилемы, ситовидных трубок и клеток-спутников флоэмы, паренхимных и других специализированных клеток. Проводящие пучки объединяют все части растительного организма, обеспечивая передвижение веществ на расстояния от десятков сантиметров до десятков метров (у древесных). Транспорт по клеточным стенкам и по цитоплазме у сосудистых растений осуществляется на небольшие расстояния, измеряемые миллиметрами, например радиальный транспорт в корнях и стеблях, передвижение веществ в мезофилле листьев.
Строение ксилемы и механизм ксилемного транспорта воды и минеральных веществ описаны в предыдущих главах (см. 5.4.4 и 6.11.2). В этом разделе будут изложены общие положения и некоторые дополнительные сведения.
 Ксилемный сок представляет собой раствор, в основном состоящий из неорганических веществ (табл. 8.1). Однако в пасоке, вытекающей из ксилемы пенька при удалении верхней части стебля, обнаружены также различные азотистые соединения (аминокислоты, амиды, алкалоиды и др.), органические кислоты, фосфорорганические эфиры, соединения, содержащие серу, некоторое количество Сахаров и многоатомных спиртов, а также фитогормоны. В ксилемном соке могут содержаться и более сложные вещества, попадающие сюда из вакуолей и цитоплазмы трахеальных элементов, заканчивающих свое развитие.
Ксилемный сок представляет собой раствор, в основном состоящий из неорганических веществ (табл. 8.1). Однако в пасоке, вытекающей из ксилемы пенька при удалении верхней части стебля, обнаружены также различные азотистые соединения (аминокислоты, амиды, алкалоиды и др.), органические кислоты, фосфорорганические эфиры, соединения, содержащие серу, некоторое количество Сахаров и многоатомных спиртов, а также фитогормоны. В ксилемном соке могут содержаться и более сложные вещества, попадающие сюда из вакуолей и цитоплазмы трахеальных элементов, заканчивающих свое развитие.
Таблица 8.1 Сравнительный состав (в ммоль/л) флоэмного и ксилемного сока у однолетнего люпина (по J. S. Pate, 1975)
| Вещества | Ксилема | Флоэма |
| Сахароза | _ | 450-470 |
| Аминокислоты | 0,7-2,6 | 13-15 |
| Калий | 2,4-4,6 | 39,0-46,0 |
| Натрий | 2,2-2,6 | 4,4-5,2 |
| Кальций | 0,4-1,8 | 0,5-1,6 |
| Магний | 0,3-1,1 | 3,5-5,8 |
| Нитраты | Следы | 0,003 |
| рН | 5,9 | 8,0 |
| 1 В мг/мл |
 Органические компоненты ксилемного сока меняются в зависимости от вида растения и от природы ионов, присутствующих в наружном растворе. Если поместить корни в солевой раствор, из которого катионы поглощаются легче, чем анионы, то количество поглощенных катионов превысит количество анионов. Растение компенсирует эту несбалансированность синтезом органических анионов, обычно карбоновых кислот: сукцината, малата, цитрата и др. В условиях, обеспечивающих высокую нитратредуктазную активность корней, важным компонентом ксилемного сока могут быть аминокислоты глутамат, аспартат, лизин и др.
Органические компоненты ксилемного сока меняются в зависимости от вида растения и от природы ионов, присутствующих в наружном растворе. Если поместить корни в солевой раствор, из которого катионы поглощаются легче, чем анионы, то количество поглощенных катионов превысит количество анионов. Растение компенсирует эту несбалансированность синтезом органических анионов, обычно карбоновых кислот: сукцината, малата, цитрата и др. В условиях, обеспечивающих высокую нитратредуктазную активность корней, важным компонентом ксилемного сока могут быть аминокислоты глутамат, аспартат, лизин и др.
Ксилемный сок по составу резко отличается от вакуолярно-го. Например, содержание иона К+ в вакуолях эпикотиля гороха достигает 55 — 78 ммоль/л, а в ксилемном соке — лишь 2 — 4 ммоль/л.
Загрузка ксилемы. Загрузка ксилемы наиболее интенсивно происходит в зоне корневых волосков. Предполагается, что в этой зоне функционируют один или два насоса. Основной насос локализован в плазмалемме клеток ризодермы и коровой паренхимы. Он обусловлен работой Н+-помп, в качестве которых выступают Н+-АТРазы и, возможно, протонперенося-щие редокс-цепи. В этой части корня катионы и анионы из клеточных стенок поступают в цитоплазму (рис. 8.1). Через клетки эндодермы с поясками Каспари вода и минеральные соли проходят только по симпласту. В паренхимных клетках пучка, непосредственно примыкающих к трахеидам или сосудам, может функционировать и второй насос, выделяющий минеральные вещества, которые через поры в стенках трахеальных элементов попадают в их полости.
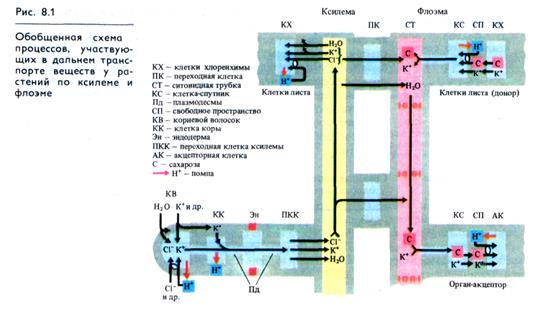
Благодаря активной работе двух насосов в трахеидах и сосудах увеличивается осмотический потенциал и, следовательно, сосущая сила. Когда вода согласно осмотическим законам поступает в полости сосудов и трахеид, в них возрастает гидростатическое давление и происходит подача жидкости в надземные части растения (корневое давление, или нижний концевой двигатель — см. 5.4.2). Восходящий ток воды и минеральных веществ, обусловленный транспирацией (верхним концевым двигателем — см. 5.4.3), осуществляется пассивно по физико-химическим законам и не сопряжен с тратой метаболической энергии, за исключением той энергии, которая необходима для поддержания открытого состояния устьиц (см. 13.6.1).
Вода и растворенные в ней вещества движутся по сосудам ксилемы с большой скоростью, иногда достигающей нескольких десятков метров в час. Скорость транспорта по трахеидам ниже, так как в этом случае жидкости приходится преодолевать первичные клеточные стенки, которые перекрывают поры в стенках продольных рядов трахеид.
Разгрузка ксилемы. Восходящий ток минеральных веществ и ряда органических соединений (амидов и аминокислот, органических кислот и др.), состав которых зависит от вида растения и условий питания, по мере продвижения по ксилеме изменяется количественно и качественно. Например, у растений кукурузы и фасоли ионы натрия, поступившие из среды в растущие участки корня, поглощаются из ксилемного сока клетками более зрелых участков корня, так что концентрация натрия в пасоке на пути к шейке корня может снизиться на два порядка. Натрий продолжает выходить из ксилемы и в тканях стебля. Радиоактивный 42К из ксилемного сока, движущегося вверх по стеблю, обнаруживается в клетках ксилемной паренхимы и коры стебля, которые поглощают также значительные количества воды. У проростков кабачка в ксилемном соке, полученном из черешков листьев (табл. 8.2), концентрация ионов калия, кальция, фосфора, нитрата более высокая, чем в соке у основания стебля. В наибольшем количестве ионы ксилемного сока поглощаются клетками листьев, о чем можно судить, например, по составу гуттационной жидкости, выделяющейся из ксилемных окончаний листа через гидатоды в условиях насыщения воздуха водяными парами. Эта жидкость (см. табл. 8.2) прошла весь путь транспорта от наружного раствора через клетки корня, по сосудам ксилемы в стебле и листе и значительно обеднена ионами по сравнению с ксилемным соком, входящим в листья.
Таблица 8.2 Концентрация ионов в ксилемной жидкости (ммоль/л) при прохождении минеральных веществ через корень, стебель и листья проростков кабачка (см. Кларксон, 1978)
| Анализируемые растворы | К + | Na + | Са2 + | Н2РО2-4 | NO-3 |
| Гуттационная жидкость | 0,5 | 0,7 | 0,9 | 0,2 | 0,4 |
| Пасока из листовых черешков | 26,2 | 0,6 | 8,0 | 0,5 | 12,9 |
| Пасока из основания стебля | 12,8 | 0,3 | 5,1 | 0,2 | 7,1 |
| Минеральная питательная среда | 0,5 | 0,2 | 0,15 | 0,01 | 1,0 |
Разгрузка ксилемы, т. е. поступление воды и ионов из сосудов ксилемы, обусловлена гидростатическим давлением в сосудах, силами транспирации и аттрагирующим действием окружающих клеток. Вода и растворенные в ней вещества через поры сосудов ксилемы попадают как в клеточные стенки (апопласт), так и в цитоплазму клеток мезофилла листа (или клеток обкладки). Поступление минеральных элементов из апо-пласта в клетки листа происходит в результате активной работы Н+-помпы (см. рис. 8.1).
Неорганические соли, различные соединения азота и фосфора, попадающие в листья с ксилемным током, необходимы для роста листьев, для поддержания их осмотического потенциала при увеличении объема клеток. Растущие листья являются мощными акцепторными зонами для компонентов ксилемного сока. Однако объем воды, доставляемой по ксилеме и испаряемой за сутки в процессе транспирации, в 5— 10 раз превосходит объем клеток листа. Поскольку вместе с водой в клетки листа транспортируются ионы, возможно перенасыщение клеток хлоренхимы солями. В тканях листьев используются по крайней мере три способа устранения избытка ионов: образование труднорастворимых осадков солей (в клеточных стенках, вакуолях, митохондриях), отток солей из листа через флоэму, а в условиях почвенного засоления — накопление и выделение солей специализированными образованиями — солевыми желёзками и волосками (см. гл. 9).
Регуляция ксилемного транспорта. Загрузка ксилемы обусловлена прежде всего функциональной активностью Н+- помпы и других ионных насосов в клеточных мембранах поглощающей зоны корня. Эта активность непосредственно связана с энергетикой дыхания и поэтому зависит от обеспеченности корня ассимилятами и кислородом. Роль фитогормонов в загрузке ксилемы изучена недостаточно.

Другой мощный регулятор ксилемного транспорта — транспирация, интенсивность которой контролируется устьичным аппаратом. Механизм устьичной регуляции обсуждается в разделах 5.4.3 и 13.6.11.
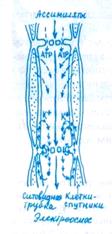
Ассимиляты из листьев поступают во флоэму, состоящую из нескольких типов клеток, специализированных в метаболи ческом и структурном отношениях. Транспортную функцию осуществляют ситовидные клетки (у низших сосудистых расте ний и голосеменных) или ситовидные трубки (у покрытосеменных растений). Паренхимные клетки-спутники выполняют энергетическую функцию; другие паренхимные клетки пучка являются передаточными или запасающими. Все эти типы клеток функционируют во взаимодействии.
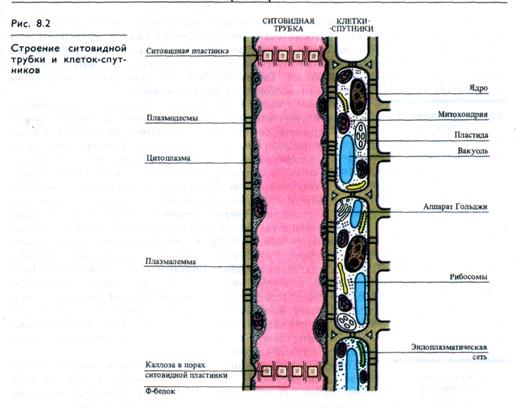
Ситовидные элементы. В ситовидных трубках плазмалемма окружает протопласт, содержащий небольшое число митохондрий и пластид, а также агранулярный эндоплазматический ре-тикулум. Тонопласт в ситовидных элементах разрушается. Зрелая ситовидная трубка лишена ядра. Поперечные клеточные стенки — ситовидные пластинки — имеют перфорации, выстланные плазмалеммой и заполненные полисахаридом калло-зой и фибриллами актиноподобного Ф-белка, которые ориентированы продольно. Ситовидные трубки многочисленными плазмодесмами связаны с клетками-спутниками (рис. 8.2).
Сопровождающие клетки. Клетки-спутники, или сопровождающие клетки, — это небольшие вытянутые вдоль ситовидных трубок паренхимные клетки с крупным ядром, элек-тронноплотной цитоплазмой, с большим количеством рибо-сом, других органелл и особенно митохондрий, обилие которых создает структуру, называемую митохондриальным ретикулумом. Клеточные стенки этих клеток пронизаны ветвящимися плазмодесмами, собранными в утолщениях стенок (плазмодесменных полях). Число плазмодесм в сопровождающих клетках в 3— 10 раз больше, чем в стенках соседних клеток мезофилла листа (например, в листе ясеня общее число плазмодесм на клетку мезофилла 800 — 3000, а в клетке-спутнике 24000). Другая особенность клеточных стенок клеток-спутников — обилие в них инвагинаций («лабиринтов»), выстланных плазмалеммой, что значительно увеличивает ее поверхность.
Организация проводящей системы в листовой пластинке. На долю проводящей системы приходится около 25% от общего объема ткани листа. Суммарная длина разветвлений проводящих пучков в 1 см2 листовой пластинки составляет в среднем от 20 до 100 см. Из этой длины на долю крупных пучков (проводящих) приходится лишь 5%, а остальные 95% — на долю ат-трагирующих тонких пучков.
Пучки, входящие в состав более толстых жилок, состоят из ксилемы и хорошо развитой флоэмы. Самые мелкие пучки включают в себя один-два трахеальных элемента, а флоэма — одну ситовидную трубку с сопровождающей клеткой. Окончания пучков во флоэмной части могут содержать только парен-химные клетки. Таким образом, первичное поступление асси-милятов в проводящую систему происходит через сопровождающие или паренхимные клетки флоэмных окончаний, из которых они перемещаются в специализированные для дальнего транспорта ситовидные элементы. У многих растений с С4- путем фотосинтеза проводящие элементы листа окружены плотно сомкнутыми клетками обкладки, отделяющими пучки от мезофилла и от межклетников. Обкладочные клетки соответствуют эндодерме осевых органов, так как в ряде случаев на радиальных (по отношению к пучку) стенках этих клеток можно видеть пояски Каспари.
 Транспорт ассимилятов в листе строго ориентирован: продукты фотосинтеза перемещаются из каждой микрозоны клеток мезофилла радиусом 70—130 мкм в сторону ближайшего к ней малого пучка и далее — по клеткам флоэмы в сторону крупной жилки. Таким образом, в состав каждой микрозоны входит всего несколько клеток мезофилла.
Транспорт ассимилятов в листе строго ориентирован: продукты фотосинтеза перемещаются из каждой микрозоны клеток мезофилла радиусом 70—130 мкм в сторону ближайшего к ней малого пучка и далее — по клеткам флоэмы в сторону крупной жилки. Таким образом, в состав каждой микрозоны входит всего несколько клеток мезофилла.
Основной транспортной формой Сахаров у большинства растений служит сахароза. В связи с этим активность инвертазы — фермента, расщепляющего сахарозу на глюкозу и фруктозу, в проводящих тканях очень низка. Концентрация сахарозы во флоэмном соке может достигать 0,8—1,0 моль/л (80 — 85% от общего сухого вещества), что в 10 — 30 раз выше, чем в клетках мезофилла. Транспортироваться по ситовидным трубкам могут и другие олигосахара (например, у ясеня — рафиноза и стахиоза), спирты (сорбит у яблони). Содержание азотистых веществ (белки, амиды, аминокислоты) во флоэмном соке не превышает 0,5%. В нем обнаружены также органические кислоты, витамины, фитогормоны (ауксин, гиббереллины, цитокинин в концентрации около 5 — 50 нмоль/л). Неорганические соли составляют 1—3% от общего количества веществ сока. Особенностью ионного состава флоэмного эксудата является высокая концентрация ионов К+ (до 50 — 200 ммоль/л) и рН порядка 8,0 — 8,5 (см. табл. 8. 1). АТР содержится в концентрации 0,4 — 0,6 ммоль/л и, по-видимому, поставляется клетками-спутниками.
Передвижение ассимилятов по ситовидным трубкам происходит со скоростью 50—100 см/ч и включает в себя три взаимосвязанных процесса: загрузку флоэмы, транспорт ассимилятов по ситовидным элементам и разгрузку флоэмы.
 Загрузка флоэмных окончаний. Всвободном пространстве клеточных стенок (в апопласте) может находиться до 20% са-харов, содержащихся в листе, куда они попадают главным образом в виде сахарозы из фотосинтезирующих клеток мезофилла. Особенности транспорта ассимилятов от мезофилла к флоэме изучены недостаточно, но, по-видимому, у разных видов растений он происходит неодинаково. Если в стенках клеток обкладки есть пояски Каспари, то сахара в этих участках должны проходить через симпласт. Многочисленные плазмо-десмы между клетками обкладки или клетками листовой паренхимы и клетками-спутниками могут способствовать сим-пластному транспорту. Развитие системы «лабиринтов» в стенках клеток, лежащих между мезофиллом и ситовидными трубками (у ряда представителей бобовых), должно способствовать транспорту ассимилятов через мембрану и апопласт-ному перемещению веществ. У некоторых растений (сахарная свекла) нет структурных приспособлений для облегчения передвижения ассимилятов. В этом случае транспорт может быть обеспечен сильно развитой системой переносчиков на мембранах клеток.
Загрузка флоэмных окончаний. Всвободном пространстве клеточных стенок (в апопласте) может находиться до 20% са-харов, содержащихся в листе, куда они попадают главным образом в виде сахарозы из фотосинтезирующих клеток мезофилла. Особенности транспорта ассимилятов от мезофилла к флоэме изучены недостаточно, но, по-видимому, у разных видов растений он происходит неодинаково. Если в стенках клеток обкладки есть пояски Каспари, то сахара в этих участках должны проходить через симпласт. Многочисленные плазмо-десмы между клетками обкладки или клетками листовой паренхимы и клетками-спутниками могут способствовать сим-пластному транспорту. Развитие системы «лабиринтов» в стенках клеток, лежащих между мезофиллом и ситовидными трубками (у ряда представителей бобовых), должно способствовать транспорту ассимилятов через мембрану и апопласт-ному перемещению веществ. У некоторых растений (сахарная свекла) нет структурных приспособлений для облегчения передвижения ассимилятов. В этом случае транспорт может быть обеспечен сильно развитой системой переносчиков на мембранах клеток.
 Измерение осмотической концентрации Сахаров показало, что в клетках мезофилла осмотическое давление ниже, чем в тонких проводящих пучках. По мере продвижения от тонких пучков к средней жилке и далее к листовому черешку общее содержание Сахаров в пучках еще более возрастает, но уже не так резко, как при переходе от мезофилла к тонким разветвлениям. Таким образом, загрузка ассимилятами проводящей системы в листе происходит против градиента концентрации, сопровождаясь затратой энергии. Источником АТР для процессов загрузки флоэмы, по-видимому, служит в 3 — 4 раза более интенсивное, чем у соседних паренхимных клеток, дыхание клеток-спутников, которые особенно богаты митохондриями.
Измерение осмотической концентрации Сахаров показало, что в клетках мезофилла осмотическое давление ниже, чем в тонких проводящих пучках. По мере продвижения от тонких пучков к средней жилке и далее к листовому черешку общее содержание Сахаров в пучках еще более возрастает, но уже не так резко, как при переходе от мезофилла к тонким разветвлениям. Таким образом, загрузка ассимилятами проводящей системы в листе происходит против градиента концентрации, сопровождаясь затратой энергии. Источником АТР для процессов загрузки флоэмы, по-видимому, служит в 3 — 4 раза более интенсивное, чем у соседних паренхимных клеток, дыхание клеток-спутников, которые особенно богаты митохондриями.
 По современным представлениям, в плазмалемме клеток-спутников (у паренхимных клеток флоэмных окончаний) функционирует Н+-помпа, направленная наружу. Закисление апо-пласта в результате работы Н +-помпы способствует отдаче ионов К+ и сахарозы клетками хлоренхимы (см. рис. 8.1). Одновременно возникший градиент рН (ΔрН) на плазмалемме клеток флоэмных окончаний приводит к поступлению в эти клетки сахарозы в симпорте с ионами Н+ (Н +-сахар — ко-транспортный механизм). Трансмембранный перенос ионов Н+ в этом случае осуществляется по концентрационному градиенту, а транспорт сахарозы — против концентрационного градиента. Этот процесс обеспечивается белками-переносчиками в плазмалемме, сродство которых к сахарам возрастает при их протонировании. Поступившие в клетки ионы Н +вновь выкачиваются Н+-помпой, работа которой сопряжена с поглощением ионов К+ (см. рис. 6.8). Сахароза и ионы К+ по многочисленным плазмодесмам переносятся в полости ситовидных трубок. Сходным образом из мезофилла в сосуды флоэмы попадают аминокислоты и другие метаболиты. Описанный механизм объясняет не только накопление сахарозы в ситовидных элементах, но и то обстоятельство, что в ситовидных трубках поддерживаются высокое содержание калия и низкая концентрация протонов. Возрастание концентрации осмотически активных веществ приводит к притоку в полость ситовидных трубок воды из окружающих тканей, прежде всего из сосудов ксилемы.
По современным представлениям, в плазмалемме клеток-спутников (у паренхимных клеток флоэмных окончаний) функционирует Н+-помпа, направленная наружу. Закисление апо-пласта в результате работы Н +-помпы способствует отдаче ионов К+ и сахарозы клетками хлоренхимы (см. рис. 8.1). Одновременно возникший градиент рН (ΔрН) на плазмалемме клеток флоэмных окончаний приводит к поступлению в эти клетки сахарозы в симпорте с ионами Н+ (Н +-сахар — ко-транспортный механизм). Трансмембранный перенос ионов Н+ в этом случае осуществляется по концентрационному градиенту, а транспорт сахарозы — против концентрационного градиента. Этот процесс обеспечивается белками-переносчиками в плазмалемме, сродство которых к сахарам возрастает при их протонировании. Поступившие в клетки ионы Н +вновь выкачиваются Н+-помпой, работа которой сопряжена с поглощением ионов К+ (см. рис. 6.8). Сахароза и ионы К+ по многочисленным плазмодесмам переносятся в полости ситовидных трубок. Сходным образом из мезофилла в сосуды флоэмы попадают аминокислоты и другие метаболиты. Описанный механизм объясняет не только накопление сахарозы в ситовидных элементах, но и то обстоятельство, что в ситовидных трубках поддерживаются высокое содержание калия и низкая концентрация протонов. Возрастание концентрации осмотически активных веществ приводит к притоку в полость ситовидных трубок воды из окружающих тканей, прежде всего из сосудов ксилемы.
Как следует из табл. 8.1, флоэмный сок содержит разнообразные ионы. С помощью радиоактивных изотопов установлено, что из ксилемного сока во флоэмный легко циркулируют калий, натрий, магний, фосфор, азот. Микроэлементы — железо, марганец, цинк, молибден — способны перемещаться по флоэме из зрелых в молодые растущие листья. Циркуляция ионов между ксилемой и флоэмой осуществляется следующим образом. В ситовидную трубку ионы могут попадать как путем транспорта из клеток мезофилла, так и не выходя за пределы листовой жилки, при участии паренхимных клеток, в клеточных стенках которых образуются многочисленные выросты цитоплазмы («лабиринты»). Клетки с таким строением сгенки, как правило, активно участвуют в транспорте ионов и называются переходными (передаточными). «Лабиринты» в стенках могут быть поляризованы. Например, у переходных клеток ксилемы стебля и листовых черешков выросты цитоплазмы есть лишь в стенке, примыкающей к сосуду, что может обеспечивать перенос ионов из ксилемного сока в симпласт ксилемной паренхимы. В переходных клетках, расположенных в жилке между сосудом ксилемы и ситовидной трубкой, выросты имеются на стенках, смежных с обоими типами транспортных систем (см. рис. 8.1). Сходный обмен ионами (и водой) может осуществляться и в стебле, где проводящие элементы ксилемы и флоэмы также сближены.
 Транспорт веществ по ситовидным трубкам.Передвижение ассимилятов по ситовидным элементам — особый случай симпластического транспорта. Механизм этого транспорта до конца не раскрыт.
Транспорт веществ по ситовидным трубкам.Передвижение ассимилятов по ситовидным элементам — особый случай симпластического транспорта. Механизм этого транспорта до конца не раскрыт.
Наиболее обоснована предложенная Э. Мюнхом (1926) гипотеза потока под давлением. Согласно этой гипотезе между фотосинтезирующими клетками листа, где в симпласте накапливается сахароза, и тканями, использующими ассимиляты (например, корнем), создается осмотический градиент, который в ситовидных трубках превращается в градиент гидростатического давления. В результате во флоэме возникает ток жидкости под давлением от листа к корню. В настоящее время эта гипотеза получила поддержку благодаря изучению механизмов загрузки и разгрузки флоэмы — решающих факторов дальнего транспорта ассимилятов. Сохраняющаяся в ситовидных трубках плазмалемма с ее свойством избирательной проницаемости является важнейшим условием, поддерживающим поток под давлением. В какой-то мере описанный механизм флоэм-ного транспорта сходен с механизмом, обеспечивающим подъем пасоки по ксилеме под действием корневого давления. Сходство особенно очевидно в случае весеннего «плача» древесных, в пасоке которых в этот период содержится много Сахаров.
Возможно, однако, что транспорт по флоэме на дальние расстояния обеспечивается не единственным механизмом. Предполагается, что движущей силой перемещения флоэмной жидкости из одной ситовидной трубки в другую через поры может быть транспорт ионов К+ в ходе электроосмотического процесса. Согласно этой гипотезе, К+ активно входит в ситовидную трубку выше ситовидной пластинки и выходит в апо-пласт в следующем членике ниже ситовидной пластинки. В результате на ситовидных пластинках возникает отрицательный электрический градиент, способствующий транспорту через них ионов К+, а с ними и молекул воды.
Предполагается также, что фибриллы актиноподобного Ф-белка в порах ситовидных пластинок во взаимодействии с миозином могут обладать контрактильными (сократительными) свойствами, что способствует передвижению жидкости по флоэме. Однако обе эти гипотезы — электроосмотическая и контрактильная — не имеют пока убедительных экспериментальных доказательств.
Разгрузка флоэмы. Для разгрузки флоэмы существенное значение имеет повышенное гидростатическое давление в ситовидных трубках. Помимо этого интенсивно растущие и запасающие органы обладают аттрагирующей силой, т. е. способны поглощать ассимиляты из флоэмы. В плазмалемме клеток акцепторной зоны функционирует Н+-помпа, которая, с одной стороны, воздействует на ситовидные трубки и клетки-спутники, закисляя апопласт и таким образом побуждая их отдавать ионы К+ и сахарозу в фазу клеточных стенок, а с другой — создает условия для поступления Сахаров и калия из апопласта в симпласт акцепторных тканей (см. рис. 8.1). Сахароза поглощается с участием мембранных переносчиков в сим-порте с протонами, а ионы К+ — по электрическому градиенту. В клетках запасающих органов, например в корнеплоде сахарной свеклы, сахароза накапливается в вакуолях. Предполагается, что через тонопласт активно переносятся ионы К+, а это способствует транспорту сахарозы. Конкретные механизмы поступления К+ и сахарозы в вакуоль неизвестны.
 Регуляция флоэмного транспорта. Транспорт ассимилятов из цитоплазмы клеток мезофилла листа (донор) к клеткам орга- нов-акцепторов, например корня, включает в себя ряд транспортных систем со своими механизмами регуляции. На уровне донора — это регуляция интенсивности фотосинтеза в хлоропластах, поступления триозофосфатов в цитоплазму и использования их на синтез сахарозы. Фотосинтез служит также источником энергии для систем активного переноса ассимилятов через мембраны (АТР для ионных насосов, субстраты для дыхания).
Регуляция флоэмного транспорта. Транспорт ассимилятов из цитоплазмы клеток мезофилла листа (донор) к клеткам орга- нов-акцепторов, например корня, включает в себя ряд транспортных систем со своими механизмами регуляции. На уровне донора — это регуляция интенсивности фотосинтеза в хлоропластах, поступления триозофосфатов в цитоплазму и использования их на синтез сахарозы. Фотосинтез служит также источником энергии для систем активного переноса ассимилятов через мембраны (АТР для ионных насосов, субстраты для дыхания).
Ключевую роль в механизме флоэмного транспорта играет загрузка флоэмных окончаний. В основе этой загрузки в клетках флоэмы лежит работа Н+-помпы, которая активируется фитогормонами, прежде всего ауксином. Абсцизовая кислота блокирует Н+/К+ обмен. Как известно, содержание АБК возрастает в тканях листа при неблагоприятных условиях, особенно при водном дефиците.
Важнейшую роль в регуляции флоэмного транспорта играет интенсивность разгрузки флоэмы в зонах потребления ассимилятов. При ослаблении аттрагирующего действия акцепторных органов снижается продольный осмотический градиент в ситовидных трубках, а следовательно, и градиент гидростатического давления — главная движущая сила флоэмного транспорта. Высокая аттрагирующая способность акцепторных органов зависит от интенсивности их роста и активности Н+-насосов. То и другое контролируется фитогормонами. Локальное воздействие абсцизовой кислоты на флоэму облегчает отдачу ассимилятов в зонах разгрузки.
Регуляция флоэмного транспорта может осуществляться и на уровне мембранных переносчиков сахарозы. Установлено, что при повышении концентрации ассимилятов в ситовидных трубках из-за интенсивного фотосинтеза или ослабления аттрагирующей активности потребляющих органов в ситовидных трубках повышается тургорное давление, что в свою очередь ингибирует работу переносчиков Сахаров. Предполагается, что при повышении тургорного давления плазмалемма в клетках флоэмных окончаний листа прижимается к клеточным стенкам, что обратимо деформирует структуру переносчика и таким образом снижает интенсивность поступления сахарозы в ситовидные трубки.
Активация транспорта ассимилятов по флоэме наблюдается при прохождении по проводящему пучку электрического импульса (потенциала действия). Флоэмный транспорт зависит от достаточного снабжения растительного организма калием, а также от температуры, так как механизм передвижения ассимилятов обусловлен работой ферментных систем (ионных насосов). Для энергетического обеспечения функциональной активности трансмембранных насосов необходим кислород.
Передвижение веществ по растению на дальние расстояния осуществляется по проводящим пучкам. По сосудам и трахеидам ксилемы вещества с водным током транспортируются от корней к верхушкам побегов. Движущие силы ксилемного транспорта — корневое давление и транспирация. Отток ассимилятов из листьев и из запасающих органов идет по ситовидным трубкам флоэмы. Загрузка как кси-лемных (в корнях), так и флоэмных окончаний (в листьях) происходит благодаря деятельности активных мембранных насосов (Н+-помп), которые функционируют в плазмалемме живых клеток, окружающих сосуды и ситовидные трубки. Вслед за поступлением осмотически активных веществ в сосуды и ситовидные трубки по осмотическим законам входит вода, и дальнейщее передвижение веществ по сосудистой системе осуществляется в результате возрастающего гидростатического давления.
 Глава 9
Глава 9

