 2015-05-26
2015-05-26 3126
3126
Рост и развитие — сложные явления, включающие не только увеличение размеров, но также процессы дифференцировки и формообразования.
Рост — это увеличение размеров тела или его частей. В его основе лежат следующие клеточные процессы: 1) увеличение числа клеток, или гиперплазия; 2) увеличение размеров клеток, или гипертрофия; 3) увеличение межклеточного вещества, или аккреция. Увеличение числа клеток есть функция клеточных делений (митоза). Увеличение размеров клеток происходит вследствие увеличения размеров функциональных внутриклеточных единиц, в первую очередь, белковой природы. Межклеточные субстанции, как органического, так и неорганического характера, связывают и аггрегируют клетки между собой в сложные комплексы.
Гиперплазия, гипертрофия и аккреция — постоянные участники ростового процесса, но их конкретный вклад зависит от «времени и места», т. е. от конкретного периода онтогенеза и характера растущей ткани. Например, число нейронов (клеток мозга) определяется уже к середине беременности, в то время как число мышечных волокон окончательно устанавливается только после рождения.
Созревание труднее поддается определению, чем рост. Часто его характеризуют как процесс перехода к зрелому состоянию, однако само понятие зрелости варьирует в зависимости от того, о каком из биологических параметров идет речь. Так, например, под половой зрелостью подразумевается полное достижение организмов функциональной способности к воспроизводству. Костная зрелость означает полную оссификацию скелета у взрослого человека. Созревание отражает темпы и сроки процесса перехода к взрослому состоянию и тесно связано с таким понятием, как биологический возраст (подробнее об этом см. в следующих подразделах). Существует значительная индивидуальная изменчивость в сроках созревания. Например, двое детей одного и того же возраста и длины тела могут различаться по такому параметру, как процент достижения ими окончательной длины тела, когда один ребенок достиг, скажем, только 65% своей дефинитивной (окончательной) длины тела, а другой — всех 80%.
Таким образом, хотя основное различие между ростом и созреванием состоит в том, что рост фокусируется на размерах тела, а созревание — на темпах их достижения, оба эти процесса неразрывно связаны.
Термины «рост» и «созревание», как правило, всегда сопровождаются еще одним понятием — «развитие». Этот термин включает в себя, по меньшей мере, два основных аспекта: биологический, когда речь идет о развитии тканей, систем органов и т. д. в процессе пренатального и постнатального онтогенеза, и поведенческий (психологический, социальный), когда ребенок осваивает навыки общения и поведения в контексте того или иного социально-культурного окружения.
К основным закономерностям роста и развития относятся необратимость, постепенность, цикличность, гетерохрония, эндогенность, индивидуальное разнообразие (Хрисанфова. Перевозчиков, 1991).
Необратимость — невозможность возвращения растущего организма к предыдущим стадиям, т. е. рост — поступательный процесс.
Постепенность — стадии роста и развития идут последовательно, одна за другой.
Гетерохрония — разные системы организма и разные признаки растут и развиваются неодновременно.
Эндогенность — отражает генетическую «заданность» ростовых процессов, удерживающих их в определенных — видоспецифических — рамках.
^ Индивидуальное разнообразие позволяет объяснить вариабельность процессов роста и созревания, реализующихся вследствие взаимодействия генетической программы и условий среды.
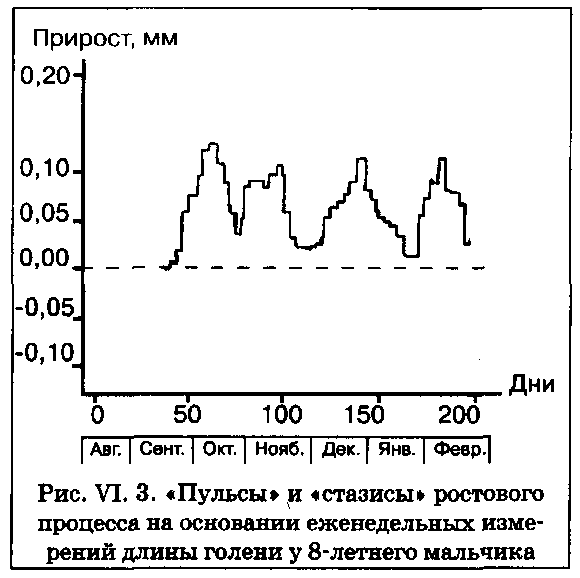
Процессы роста и развития характеризуются цикличностью, т. е. происходят не плавно и постепенно, а циклами, или «пульсами». Многочисленными исследованиями последних лет показано, что периоды активного роста перемежаются с периодами его торможения. Механизм чередования этих фаз можно связать с чередованием процессов гиперплазии и гипертрофии. Какова частота ростовых циклов, или пульсов? На рис. VI. 3 изображена кривая роста длины голени 8-летнего мальчика, измерявшегося еженедельно на протяжении нескольких месяцев. Как видно, периоды активного роста разной степени интенсивности чередуются с периодами затухания или почти полного его отсутствия — так называемыми «стазисами». Понятно, что выявление пульсов и стазисов требует проведения особенно тщательных измерений и новых методических подходах. Благодаря изобретению нового измерительного прибора, названного «кнемометром» (от греческого «кнемо» —голень), стало возможным исследовать не только недельные, но даже суточные, ритмы. Пульсы и стазисы различных сегментов тела не совпадают по времени — классический пример гетерохронии. В одном японском исследовании при ежедневных измерениях двух детей — 7,5 и 6 лет — было показано, что рост в длину происходил в основном за счет нижних конечностей, за исключением сезонных «пульсов» роста туловища в весеннее время (еще один убедительный пример гетерохронии). Сезонные циклы ростовых процессов выявлены в работах многих исследователей, но мы еще вернемся к ним в подразделе, посвященном средовым факторам, оказывающим влияние на рост.
Одним из основных понятий ауксологии и возрастной антропологии в целом является понятие «онтогенеза». Этот термин был введен в биологию Э. Геккелем при формулировании им биогенетического закона. Онтогенез, отражающий изменения в процессе индивидуального развития организмов (от греч. «онтос» — бытие), противопоставлялся филогенезу — изменениям таксономических единиц в процессе эволюции (от греч. филон — раса), и относился вначале только к периоду развития. Однако впоследствии под онтогенезом стали понимать все последовательные преобразования организма, от зачатия до окончания жизненного цикла.
Постнатальный онтогенез человека и методы его изучения
Возрастная периодизация постнатального онтогенеза человека
Существует немало периодизаций онтогенеза, древнейшие из которых принадлежат еще античным ученым. В последующие столетия в разработку периодизации индивидуального развития внесли свой вклад многие ученые (например, в XX в. — немецкие ученые С. Штрац, Г. Гримм, российские ученые — В.В. Бунак, И.А. Аршавский и др.) В нашей стране наиболее распространенной считается схема возрастной периодизации постнатального развития человека, принятая на VII Всесоюзной конференции по проблемам возрастной морфологии, физиологии и биохимии в 1965 г. (табл. VI. 1).
Таблица VI. 1.
^ Схема возрастной периодизации постнатального онтогенеза человека

Существуют и другие схемы периодизации. Так, например, в книге немецкого ученого X. Ремшмидта (1994) приводятся следующие возрастные стадии детства и взросления:
Младенчество — от рождения до конца 1-го года жизни.
Детство — от рождения до конца 13-го года жизни.
Подростковый возраст — 12/14-18 лет.
Юность — 18-21 год.
Одна из последних попыток периодизации онтогенеза принадлежит известному американскому антропологу и ауксологу Б. Богину (Bogin, 1999). Основные стадии жизненного цикла человека, по Б. Богину, с краткими характеристиками представлены в табл. VI. 2.
ТаблицаУ!. 2. Стадии жизненного цикла человека (Bogin, 1999)

Предлагая новую периодизацию, Б. Богин дает новые трактовки выделенным стадиям, с позиций эволюционной антропологии,подчеркивая особую эволюционную значимость в развитии человека таких стадий, как детство, ювенильный и подростковый периоды. Можно соглашаться или не соглашаться с предлагаемой Богиным схемой периодизации онтогенеза, однако представляется весьма уместным рассмотреть те особенности индивидуального развития человека, которые отличают его от остальных животных, в том числе, и от приматов. Сразу же отметим, что мы будем рассматривать в дальнейшем только период постнатального онтогенеза, причем те его стадии, которые касаются роста и развития.
Индивидуальное развитие человека существенно отличается от онтогенеза животных. По некоторым параметрам человек сближается с приматами, хотя опять-таки велики и отличия между ними.
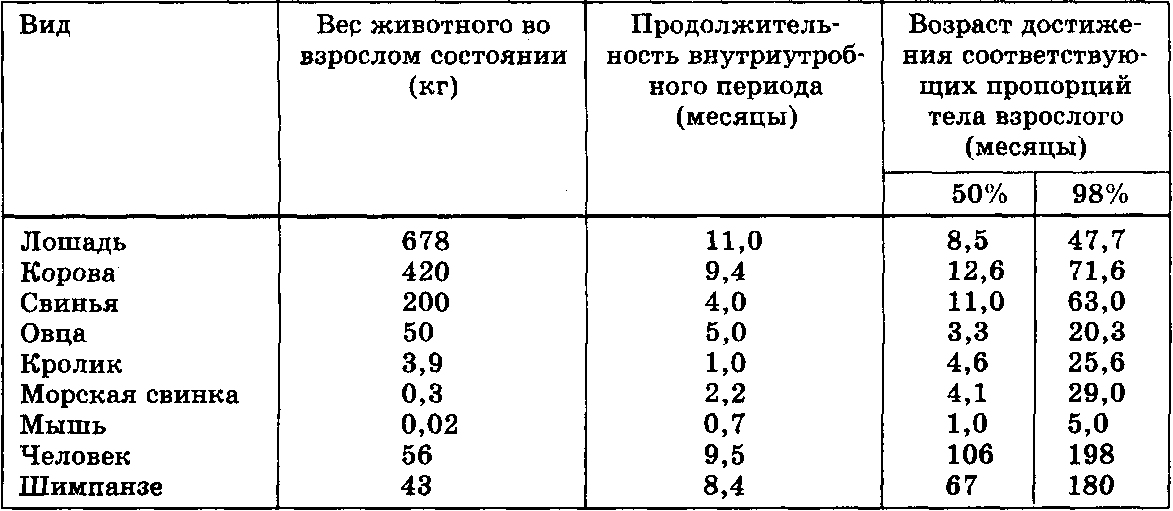
Представители семейства млекопитающих сильно варьируют как по показателям продолжительности жизни, так и по размерам тела. Исключая приматов (вместе с человеком), в ряду млекопитающих наблюдается закономерность удлинения отдельных периодов онтогенеза с увеличением размеров тела. Это в первую очередь справедливо для соотношения между весом тела взрослой особи и продолжительностью внутриутробного развития, а также, хотя и в меньшей степени, —возраста достижения 50% или 98% дефинитивного (окончательного) веса тела. При включении в анализ соответствующих данных по приматам, общая картина несколько усложняется (табл. VI. 3; табл. VI. 4). Человек живет дольше других приматов — свыше 70 лет, тогда как максимальная продолжительность жизни шимпанзе в неволе составляет около 45 лет. Удлинение общей продолжительности жизни означает и удлинение отдельных ее фаз, или периодов.
Таблица VI. 3.
Продолжительность ростового периода у млекопитающих, % к продолжительности жизни
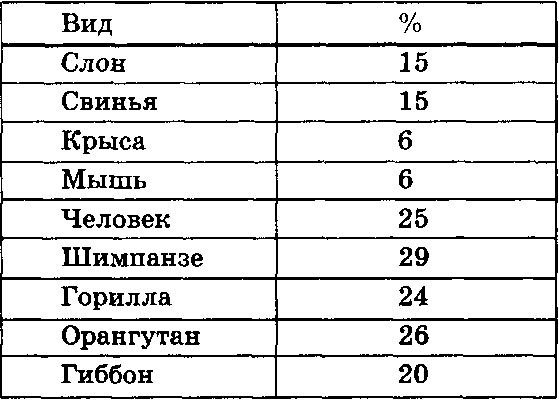
Таблица VI. 4.
Некоторые характеристики процессов роста у представителей различных видов млекопитающих (самки)
^ Сравнительный онтогенез человека и животных
Важнейшей чертой человека, приобретенной в процессе эволюции, является удлинение периода, отделяющего момент рождения от половозрелости, т. е. от взрослого состояния. Удлинение периода детства, сравнительно с остальными млекопитающими, включая приматов, достигает у человека максимального выражения.
При этом для большинства млекопитающих характерен постепенный переход от детского к взрослому состоянию, без драматического увеличения скоростей роста, как это наблюдается у человека. У млекопитающих с высоким уровнем социальной организации, например, у волков, диких собак, львов, слонов и, конечно, приматов, можно выделить специальный, «ювенильный» период, отделяющий детскую стадию от половозрелости, однако и он, как правило, проходит на фоне снижения скоростей роста.
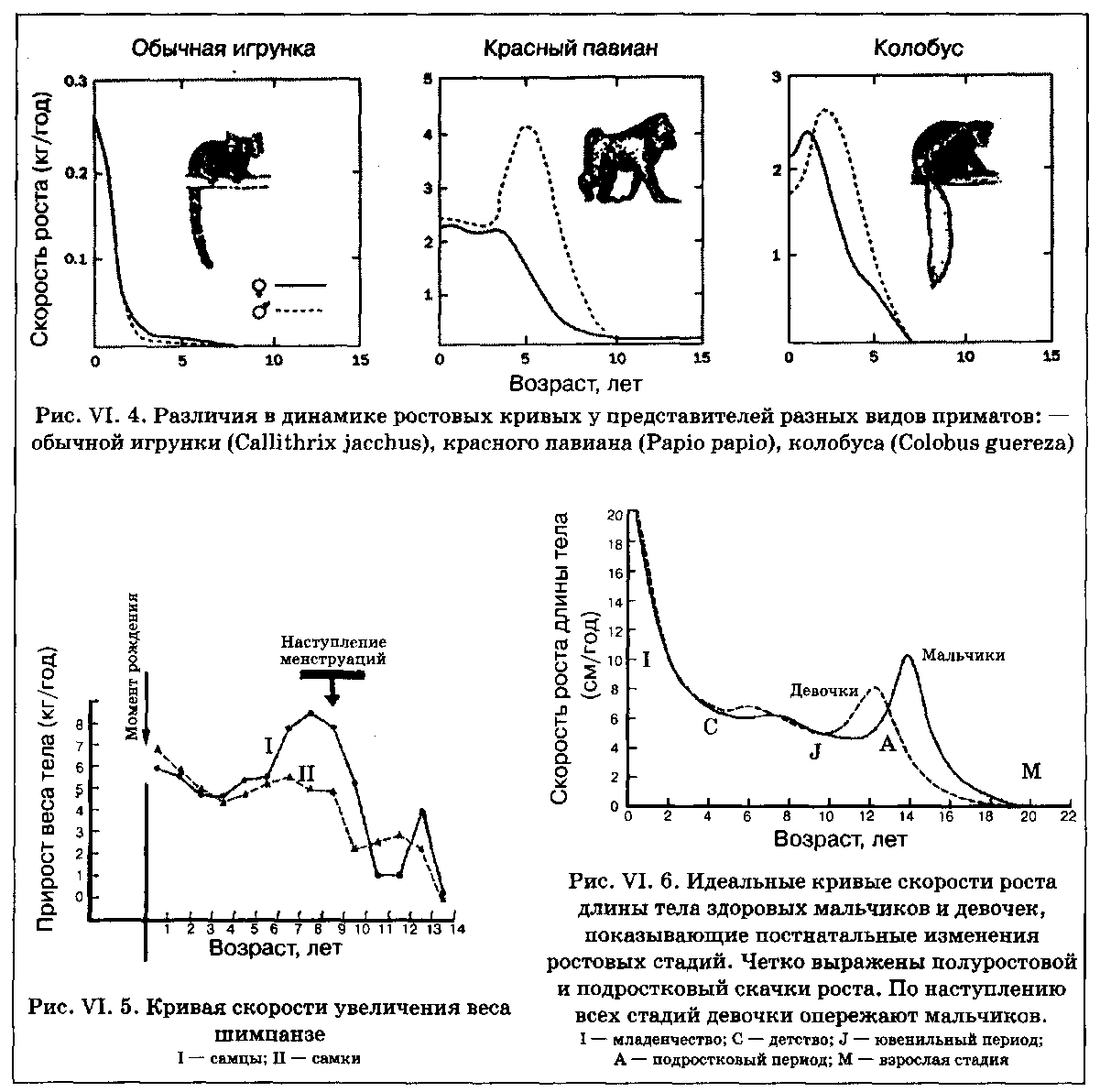
Характер постнатального роста приматов характеризуется чрезвычайным разнообразием. На рис. VI. 4 изображены кривые ежегодных приростов веса тела (скоростей роста) у представителей различных видов приматов. Как видно из рисунков, пубертатное увеличение скоростей роста (ростовой спурт) полностью отсутствует у мармозеток, обнаруживает значительный половой диморфизм (сильная выраженность у самцов и практическое отсутствие у самок) среди павианов и отчетливую выраженность для обоих полов у колобусов.
У шимпанзе ход кривых возрастного увеличения веса тела (рис. VI. 5) явно напоминает таковой у человека (рис. VI. 6). В обоих случаях значительные скорости роста при рождении постепенно снижаются до определенного предела, а затем вновь резко увеличиваются, характеризуя наступление подросткового скачка роста. Таким образом, у человекообразных обезьян наблюдается не только тенденция к увеличению продолжительности детства (свойственная всем приматам), но и проявляется отсрочка времени наступления полового созревания. Этот вывод, основывающийся, как уже отмечалось, на возрастных изменениях веса тела, кажется не столь обоснованным, когда речь идет о скелетных размерах. По мнению Б. Богина, отсрочка времени наступления полового созревания у человека выражена намного сильнее, чем у других приматов, включая и человекообразных обезьян. Эта особенность, наряду со значительно большей интенсивностью роста во время пубертатного периода, приводит к появлению уникальных видоспецифических для человека ростовых кривых.
В табл. VI. 5 суммирована информация о некоторых событиях пубертатного периода у макак резусов и шимпанзе. Как видно из таблицы, у шимпанзе, по сравнению с макаками, происходит значительная отсрочка событий пубертатного периода, как у самцов, так и у самок. При этом самки представителей обоих видов приматов опережали самцов по всем перечисленным показателям. При анализе данных, приведенных в таблице, необходимо подчеркнуть, что у обезьян, по сравнению с человеком, наблюдается иная последовательность событий пубертатного периода: у человека наступление половой зрелости (менархе у девушек) отмечается после прохождения пика максимальной скорости роста. У обезьян эти соотношения нарушаются: у макак резусов половозрелость у самок отмечена даже до начала спурта, а у самок шимпанзе — в среднем до начала пубертатного скачка роста.
Таблица VI. 5. Возраст подросткового скачка роста и половозрелости у макак резусов и шимпанзе
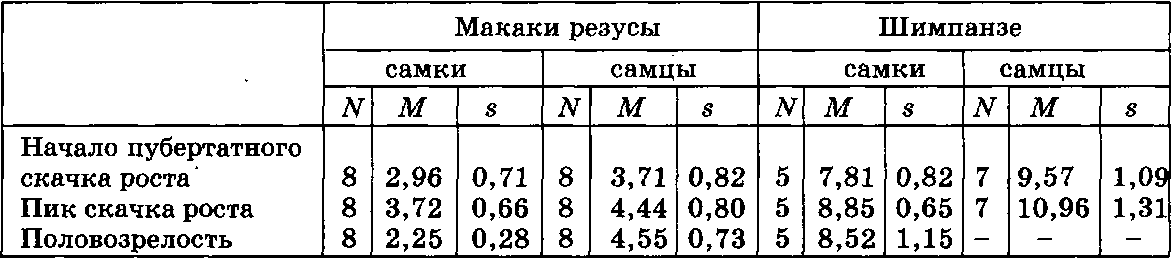
Примечание. N — число особей; М — средняя величина; s — среднее квадратическое отклонение.
Удлинение детства и замедление роста и развития расширяют возможности обучения и приобретения социальных навыков. Наметившись у приматов, эта тенденция получает свое окончательное развитие у человека. По сравнению с человекообразными обезьянами, у человека позже прорезывается первый моляр, позже начинается менструирование и наступает период деторождения. Очевидно, с эволюционной точки зрения, развитие вышеупомянутых особенностей, наряду с развитием мозга, языка и культуры, дало человеку значительные преимущества, связанные, в первую очередь, с освоением социально-культурного опыта. Продолжительное детство, согласно классическому объяснению, дает человеку сразу несколько преимуществ, главнейшими из которых являются более длительный период роста мозга; наличие большего времени для освоения технических навыков, например, для изготовления орудий; больше времени для социализации, игры, для развития системы социальных ролей и культурного поведения.
Б. Богин в своих последних работах дает еще одно объяснение тому, почему для человека как биологического вида характерен столь продолжительный период детства. По мнению Богина, небольшие размеры тела и подчеркнутая «детскость», выражающаяся прежде всего в соотношениях мозгового и лицевого черепа, могли служить своеобразными стимуляторами родительских чувств со стороны взрослых членов сообщества. В подтверждение своей гипотезы Богин приводит данные о том, что взрослые индивиды в большей степени склонны защищать и кормить малышей с выраженной «неотенией» лицевых черт. При сравнении детей, подвергавшихся насилию со стороны взрослых, и контрольной группы выяснилось, что у первых было в среднем меньшее соотношение между мозговым и лицевым черепом, т. е. они были менее «неотеничны», чем дети из контрольной группы, о которых взрослые нежно заботились.
В этой связи интересно проанализировать, как менялось соотношение между отдельными стадиями развития в процессе эволюции у представителей различных групп гоминид. На рис. VI. 7 представлено соотношение между отдельными стадиями онтогенеза у некоторых представителей гоминид. Как видно, в процессе эволюции увеличивалась продолжительность всех стадий, в особенности касающихся ростового периода. Конечно, изучение ростовых процессов у ископаемых гоминид неизбежно связано с тем, что соответствующие хронологические маркеры могут быть воспроизведены только по аналогии с современными популяциями (подробно см. раздел II). Это значительно усложняет задачу исследователей. Периодизация онтогенеза ископаемых гоминид базируется на особенностях зубной и костной системы. Существуют некоторые соотношения между временем прорезывания первых постоянных моляров (коренных зубов) и размерами мозга. Так, например, у австралопитековых (объем мозга 300-400 см3) первые моляры, вероятно, прорезывались в возрасте от 3 до 3 с половиной лет. Продолжительность жизни этих гоминид насчитывала, скорее всего, 35-40 лет. У Homohabilis с объемом мозга 580-750 см3 прорезывание первого моляра происходило примерно в 4,5 года, а продолжительность жизни составляла около 50 лет и т. д. У Homo erectus (объем мозга — 900-1100 см3) появление первых моляров отмечалось, вероятно, в 5 с половиной лет. Люди современного типа (Homo sapiens), обладающие самым крупным мозгом (в среднем объем мозга — 1400 см3), также отличаются наиболее длительным периодом прорезывания первых моляров, которые появляются у них около 6 лет. В то же время современные люди отличаются наибольшей для всех приматов продолжительностью жизни и ростовых фаз онтогенеза (рис. VI. 9).

Методы изучения постчатального роста человека
Основным методом изучения постнатального соматического роста остается антропометрический (от греч. «антропос» — человек, «мет-рос» — измерения), который дает возможность изучения изменений размеров тела с возрастом. Существуют два основных «тактических» приема в изучении ростовых процессов. Это уже упоминавшиеся «поперечные» и «продольные» (лонгиту-диналъные) исследования.
При поперечном исследовании делается как бы поперечный срез популяции (отсюда и название), когда измеряется большое число детей различных возрастов, но каждый индивид попадает в общую выборку только один раз.
При продольном исследовании одни и те же дети измеряются последовательно в каждом возрасте с теми или иными, обычно регулярными интервалами, на протяжении определенного отрезка времени — в зависимости от стратегии исследовательского проекта.
Оба метода находят широкое применение в ауксологии. Результаты поперечных исследований используются в основном для составления популяционных стандартов роста, при сравнительных исследованиях сопоставления характера роста в зависимости от воздействия тех или иных факторов внешней среды. С другой стороны, информацию об истинных скоростях роста можно получить только при анализе продольных обследований.
Иногда встречаются исследования «третьего рода», так называемые «смешанно-продольные», когда несколько исходных возрастных когорт изучаются продольно в течение ограниченного отрезка времени, с последующим их пересечением. Допустим, например, что в 1990 г. были впервые обследованы две группы детей 7 и 12 лет. В дальнейшем их измеряли повторно (лонгитудинально), в течение 5 лет, чтобы получить два отрезка ростовой кривой: с 7 до 12 и с 12 до 17 лет. Кажущимся преимуществом этого метода является его экономичность в смысле затрат времени: на сбор «истинно» продольной информации об этом временном периоде ушло бы еще 5 лет. Однако недостатки метода значительно более существенны: претендуя на «продольность», он дает лишь очень приблизительную характеристику скорости роста, вследствие отсутствия точной информации в точке (точках) перекреста. По крайней мере, опробовав эту тактику в одном из наших проектов, мы впоследствии отказались от нее, в частности, потому, что значительные диахронные изменения существенно исказили истинную картину ростовых процессов в популяции (Миклашевская, Соловьева, Година,1988).
Выбор метода обследования целиком зависит от поставленной задачи: для того, чтобы дать характеристику статуса физического развития растущей части той или иной популяции, вполне достаточно добросовестно выполненного поперечного обследования. Если же целью исследования является мониторинг тех или иных ростовых закономерностей, например, проявления сезонных изменений и т. д., идеальным методом будет продольное обследование.
Изучение постнатального соматического роста, как уже говорилось, осуществляется в основном с помощью антропометрических (измерительных) и антропоскопических (описательных) методик. Вкратце упомянем, какие именно измерения и приборы используются в ауксологии (некоторые описательные — антропоскопические — методики приведены в разделе, посвященном биологическому возрасту).
Прежде всего отметим, что число измерений, проводимых на живом человеке, может быть практически бесконечным, поэтому выбор конкретной измерительной программы всегда зависит от задач исследования и возможностей исследовательской группы. Наиболее широко распространены измерения тотальных размеров — длины и веса тела. Длина тела, а также его пропорции (длина туловища, длины конечностей, их сегментов и т. д.) измеряются, как правило, антропомером (рис. VI. 8). При этом должна быть тщательно соблюдена поза, в которой стоит измеряемый: прямо, но без особого напряжения, ступни по возможности соприкасаются пятками (исключение составляют индивиды с выраженной Х-образной формой ног), расстояние между носками — 10-15 см, спина выпрямлена; грудь слегка выдается вперед; живот подобран; руки выпрямлены; пальцы прижаты к телу; плечи находятся в естественном положении— они не должны быть чрезмерно опущены или подняты, вытянуты вперед или отведены назад; голова ориентирована так, чтобы глазнично-ушная горизонталь (линия, проходящая через козелок уха и наружный край глазницы) была параллельна полу. Измеряемый, раздетый до трусов, должен стоять спокойно и не менять позу в течение всего процесса измерений. Необходимо также учитывать суточные изменения длины тела: за счет уплощения межпозвоночных дисков она может уменьшаться к вечеру.
Мы специально так подробно остановились на описании процедуры измерений только длиннотных размеров тела. Зачастую приходится сталкиваться с пренебрежительным отношением к антропометрии. В наш век компьютеров и молекулярной биологии антропометрическая методика кажется простой, легкой и не требующей особой подготовки. Однако это обманчивая простота: существует множество методических рекомендаций и ограничений, направленных на получение достоверных и воспроизводимых результатов. Без точного соблюдения этих рекомендаций измерения превращаются в пустую трату времени, а полученная в итоге информация — в набор бессмысленных цифр. Не имея возможности дать подробную характеристику всем методическим рекомендациям, мы надеемся в ближайшем будущем осуществить издание специального методического пособия, так как необходимость в нем давно назрела (фактически со времен классического труда В.В. Бунака «Антропометрия», не переиздававшегося с 1941 г. и давно ставшего библиографической редкостью, аналогичных пособий в свет не выходило).
Следующая группа измерительных признаков, основывающихся на скелетных размерах, — диаметры тела: плеч, таза, поперечный и продольный диаметры грудной клетки. Они измеряются специальным инструментом — большим толстотным циркулем. Нередко для учета роста костного компонента измеряются также диаметры суставных мыщелков — локтя, запястья, колена и лодыжки — с использованием скользящего циркуля.
Информацию о развитии мягких тканей, в первую очередь мышц, можно получить при измерении окружностей, или обхватов, грудной клетки, плеча, предплечья, бедра, голени и т. д. Измерения производятся сантиметровой лентой и также требуют особой тщательности и четкой стандартизации.
^ Жировые складки на туловище и конечностях измеряются калипером, обеспечивающим стандартный нажим на мягкие ткани. По величи-
не жировых складок и окружностей тела рассчитываются жировой и мышечный компоненты тела (см. раздел, посвященный составу тела).
Большая группа измерений производится на голове и лице. Их особая ценность в том, что информацию можно получить уже в раннем детстве (например, об окружности головы): рост головы (как и мозга) заканчивается раньше других частей тела.
В принципе, число измерений может быть практически бесконечным. В связи со специальными задачами проводятся измерения разных частей тела, например, измерения стопы, кисти для целей легкой промышленности и составляющих их элементов.
Подытоживая сказанное выше, необходимо отметить, что длина и вес тела обеспечивают информацию о росте ребенка в целом, а остальные размеры — о росте отдельных частей и тканей тела. Костные диаметры описывают общие размеры скелета, обхваты конечностей дают представление о развитии мышечной ткани, жировые складки — о количестве и распределении подкожного жира. Дети, имеющие одинаковые длину и даже вес тела, могут сильно различаться по пропорциям, форме тела и развитию мягких тканей.
Измерительные признаки служат основой для расчета и вычисления ряда производных признаков, а также соотношений размеров — индексов. Хотя существует огромное количество различных индексов*, мы упомянем здесь только один из них:
* Метод индексов неоднократно подвергался справедливой критике, в частности, потому, что составные части индекса всегда оказываются высоко скоррелированными.
индекс массы тела (ИМТ), или индекс Кетле. Это один из многочисленных весоростовых индексов, расчет которого производится по формуле P/L2, где Р — вес тела, L — длина тела. В последнее десятилетие этот индекс получил широкое распространение в ростовых исследованиях и вошел во все учебники по ауксологии, изданные, в основном, на Западе. При простоте расчета он дает хорошее представление о компонентах массы тела, в первую очередь о развитии жирового слоя, хорошо согласуется с данными о питании (или недоедании) детей и является составной частью различных популяционных стандартов (см. раздел «Процентильный метод»).
Основные характеристики постнатального роста
Как уже говорилось, ростовые графики бывают двух видов: одни отражают постоянное увеличение длины тела с возрастом — это так называемые «дистантные» кривые, другие — кривые приростов, или «скоростные», характеризуют скорость ростового процесса и намного лучше «дис-тантных» отражают его суть.
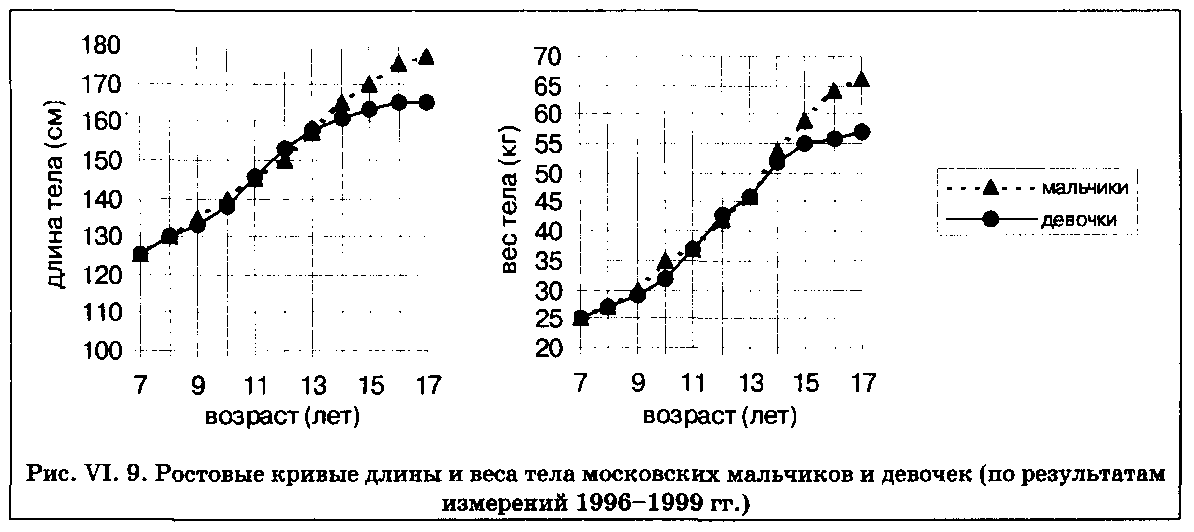
На рис. VI. 9 изображены кривые, отражающие рост длины и веса тела в разных возрастно-половых группах московских мальчиков и девочек от 7 до 17 лет. Согласно общепринятым в ауксологии правилам, возрастная группировка должна производиться таким образом, чтобы середина возрастного интервала (а значит, и средний возраст группы) приходилась на целое число лет. Так, в группу 7-летних входят дети от 6 лет 6 мес. до 7 лет 5 мес. 29 дней (средний возраст 7 лет), в группу 8-летних — дети от
7 лет 6 мес. до 8 лет 5 мес. 29 дней и т. д. Кривые, представленные на рис. VI. 9, демонстрируют собой типичный пример дистантных кривых. Они отражают размеры тела, достигнутые детьми в том или ином возрасте, а также «расстояние», пройденное ими до завершения пути, т. е. до взрослого состояния. Дистантные кривые, по определению, являются выровненными, так как на них наносятся усредненные значения признаков для большого количества детей, обследованных в том или ином возрастном интервале. При этом значительная часть информации относительно индивидуальных вариаций ростовых характеристик оказывается «растворенной» в масштабе этих усредненных кривых.
Дистантные кривые обычно используются для составления ростовых стандартов, или оценочных таблиц. Для этого проводятся в основном поперечные обследования здоровых детей в тех или иных группах населения, с последующей разработкой стандартов теми или иными статистическими методами. Наиболее распространены в настоящее время так называемые «процентильные, или пер-центильные, стандарты» (см. раздел «Процентильный метод»). Необходимо помнить, что стандарты, или оценочные таблицы (последний термин используется в последнее время гораздо чаще, особенно на Западе, в силу того, что в нем менее выражен подтекст обязательного соответствия некоему «эталону») должны быть получены на репрезентативном материале и удовлетворять, по крайней мере, двум требованиям — месту и времени, т. е. должны строиться для конкретной популяции на определенном отрезке времени. Эти два требования объясняются тем, что ростовые характеристики находятся в зависимости от определенных факторов среды и со временем могут меняться (см. соответствующие разделы).
Кривые ростовых прибавок, или скоростей роста, имеют совершенно иную форму. Для их построения необходимо иметь продольные (лонгитудинальные) данные, так как только они отражают истинный характер ростовых процессов в популяции. На рис. VI. 6. изображены кривые скорости роста длины тела для девочек и мальчиков в возрасте от 0 до 18 лет. График показывает, что скорость роста длины тела неуклонно снижается сразу же после рождения. Ребенок растет и становится больше, но процесс этот протекает более медленно, и погодовые (полугодовые, помесячные и т. д.) прибавки уменьшаются. Иногда, как это видно на рисунке, отмечается так называемый «полуростовой» скачок увеличения скорости у детей 7-8-летнего возраста. Скорость роста достигает своего минимума как раз перед началом пубертатного спурта (ускорения роста, подросткового скачка роста), после чего начинает резко возрастать. В среднем увеличение скоростей роста у девочек начинается на два года раньше, чем у мальчиков. Девочки в среднем раньше заканчивают рост, примерно к 16 годам, в то время как мальчики продолжают расти до 18-19 лет, однако у значительного числа индивидов рост продолжается: у девушек — после 18 лет, у юношей — и после 20 лет.
При внимательном рассмотрении ростовых кривых становится понятным, за счет чего возникают половые различия (половой диморфизм) в длине (и других размерных признаков) тела взрослых мужчин и женщин. Как видно на рис. VI. 9, вплоть до наступления пубертатного периода различия в росте между мальчиками и девочками очень невелики. Хотя девочки первыми вступают в пубертатный период и максимальная скорость роста у них примерно на 2 см меньше, чем у мальчиков (см. рис. VI. 6), основные различия размерных признаков формируются после 16 лет, т. е. после того, как девочки перестают расти. Мальчики за счет более длительного периода предпубертатного роста, более интенсивного ростового скачка и более продолжительного периода постпубертатного роста в среднем обгоняют девочек по длине тела на 10 см.
Попутно отметим, что два перекреста ростовых кривых (см. рис. VI. 9) — первый, когда девочки обгоняют мальчиков за счет более раннего вступления в пубертатный период, и второй, когда мальчики обгоняют девочек после окончания у последних ростового периода, — также служат хорошими «маркерами», отражающими влияние условий среды. В некоторых популяциях в целом типичная картина наличия двух ростовых перекрестов может искажаться под влиянием тех или иных условий жизни (см., например, раздел «Рост в высокогорье»).
Помимо отчетливо выраженного пубертатного скачка роста, у части детей выявляется так называемый «полуростовой скачок» — небольшое увеличение скоростей роста в 6-7-летнем возрасте, т. е. до наступления «основного» скачка. У девочек полуростовой скачок наступает в среднем раньше, чем у мальчиков, но обнаруживается он чаще у мальчиков, вероятно, вследствие большей интенсивности приростов у последних.
Полуростовой скачок удается обнаружить не всегда, причем, не только по объектным причинам (он отсутствует или очень мал), но и вследствие некоторых методических просчетов: годовой интервал (с которым обычно и проводятся измерения) может быть слишком велик для обнаружения этого, в общем, не столь значительного увеличения скоростей роста.
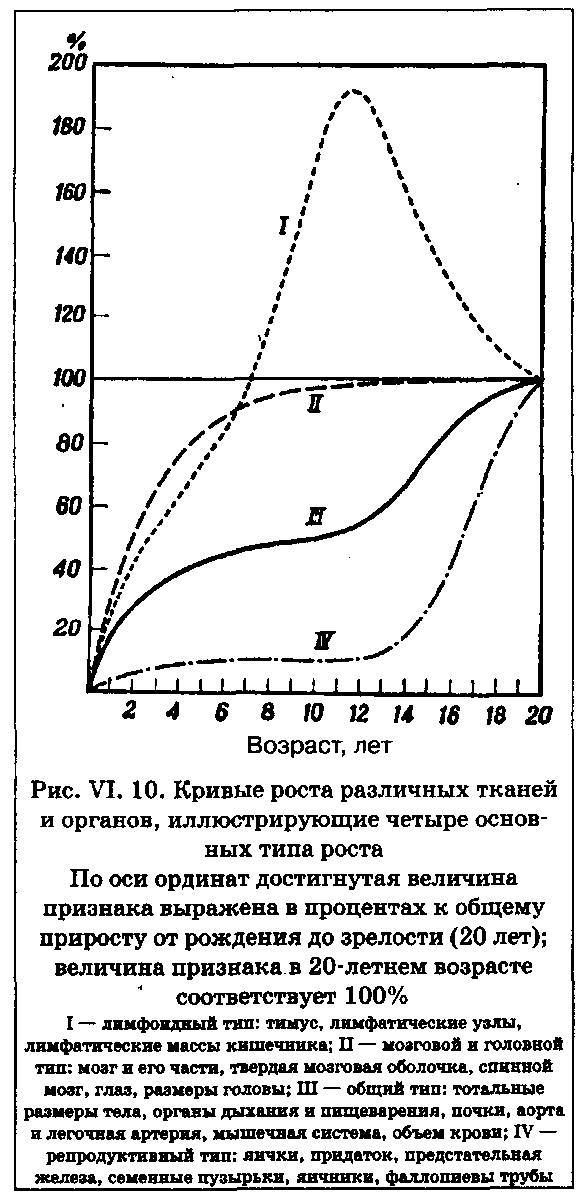
Кривая роста тела в длину, представленная на рис. VI. 9, отражает лишь один из возможных вариантов постнатальных возрастных изменений, и ей соответствуют далеко не все ткани тела. На рис. VI. 10 изображены кривые роста различных частей и тканей тела. При этом «общий» тип кривой соответствует возрастным изменениям скелетных размеров и веса тела, кривая роста мозга отражает увеличение массы мозга, а «репродуктивная» кривая — вес гонад и других половых органов. Хорошо видно, что рост мозга заканчивается значительно раньше, чем рост других органов и тканей. Во внутриутробном периоде он растет намного быстрее других частей тела, так что мозг новорожденного ребенка в среднем соответствует 25% своего окончательного размера. Еще через 5 лет масса мозга достигает 90% дефинитивной величины, а к 7-9 годам по размерам практически не отличается от мозга взрослого. В динамике возрастных изменений мозга фактически не наблюдается ростового скачка. Некоторый, хотя и довольно незначительный, скачок роста отмечен для продольного и поперечного диаметров головы, но это скорее связано с утолщением костей черепа, развитием синусов и т. д., нежели с ростом самого мозга.
Рост лицевых размеров в целом следует «общему» типу. Их возрастная динамика в среднем имеет выраженный ростовой скачок, особенно заметный у признаков нижней челюсти: подбородок становится длиннее и больше выступает вперед.
Рост лимфоидной ткани (рис. VI. 10) достигает максимума в предпубертатном периоде, а затем довольно резко снижается до «взрослых» значений. Репродуктивные органы, напротив, отличаются замедленным ростом до наступления подросткового периода и последующим бурным спуртом.
Рост большинства размерных признаков тела, за исключением показателей жироотложения, следует «общему», описанному выше, типу, т. е. протекает с наибольшей скоростью в младенчестве и раннем детстве, затем несколько замедляется, вновь увеличивается в подростковом периоде и в конце концов завершается по достижении размеров, свойственных взрослому. Рост многих признаков продолжается и в третьей декаде жизни.
При общем сходстве ростовых закономерностей, между отдельными признаками имеются различия по величине подросткового скачка роста и времени его наступления. Так, сравнение графиков роста длины туловища и длины ноги показывает, что в детстве и предпубертатном периоде конечности растут быстрее туловища и раньше достигают пубертатного спурта. Таким способом достигаются различия в пропорциях тела. Соотношение длины туловища к длине ноги у 10-11-летних девочек и мальчиков примерно одинаково, впоследствии мальчики опережают девочек по длине ноги, и эти различия сохраняются вплоть до взрослого состояния.
Аналогичные изменения происходят и в соотношении: ширина плеч/ ширина таза. Отчетливое увеличение ширины плеч по отношению к ширине таза является характерным признаком пубертатного периода мальчиков, тогда как женскому полу свойственны противоположные соотношения. Следует особо подчеркнуть важность относительных показателей: по абсолютным значениям различия, к примеру, в ширине таза, могут быть невелики, но относительно ширины плеч девочки практически всегда обгоняют мальчиков на протяжении ростового периода от 6 до 17 лет. Подобная гетерохрония, как уже говорилось, служит основным источником формирования показателей полового диморфизма, отличающего взрослых представителей вида Homo sapiens.
Возрастные изменения жировой ткани отличаются несколько иными закономерностями. Измерения показателей жироотложения могут осуществляться с помощью калиперометрии (см. выше) или ультразвукового метода. При этом скорость роста жировой ткани после рождения продолжает у младенцев увеличиваться примерно до 9-месячного возраста (в среднем), затем несколько снижается, и в 6-8 лет начинает вновь увеличиваться. Девочки в среднем характеризуются большим количеством подкожного жира, чем мальчики, причем, эти различия усиливаются с возрастом. Начиная примерно с 8 лет, наблюдаются отчетливые половые различия в ходе кривых роста жировой ткани для мальчиков и девочек. У мальчиков толщина жировых складок на конечностях снижается, рост жирового слоя на туловище несколько стабилизируется. У девочек развитие жира на конечностях несколько приостанавливается, но не снижается, а жир на туловище продолжает расти.

