 2015-05-26
2015-05-26 2077
2077
Иммунная система включает специализированную, анатомически обособленную лимфоидную ткань, «разбросанную» по всему организму в виде различных лимфоидных образований и отдельных клеток.
Различают первичные - центральные (костный мозг и тимус) и вторичные - периферические (селезенка, лимфатические узлы, скопления лимфоидной ткани) органы иммунной системы. Все они взаимосвязаны системой кровообращения, лимфотока и единой системой иммунорегуляции.
Первичные - центральные органы иммунной системы
Центральные органы иммунной системы - костный мозг и тимус выполняют важнейшие функции, обеспечивая самообновление иммунной системы, в этих органах идут процессы пролиферации клеток предшественников, их дифференцировка и созревание, вплоть до выхода в циркуляцию и заселения периферических органов иммунной системы зрелыми иммунокомпетентными клетками.
Рис. 4. Клетки крови формируются в костном мозге (фотография).
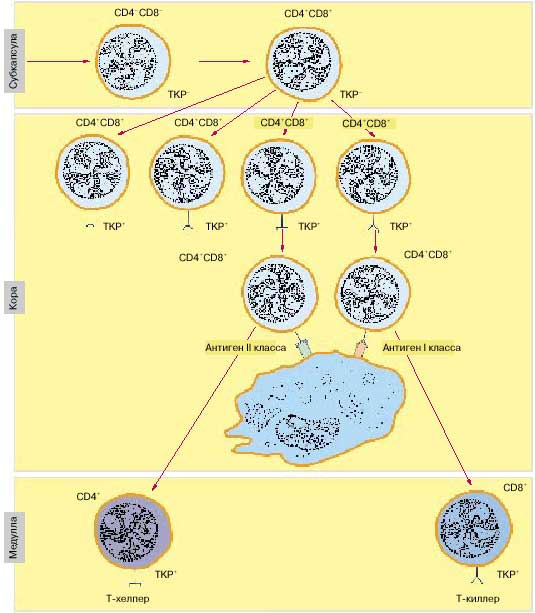
Рис. 5. В тимусе происходит созревание Т-хелперов и Т-киллеров, которые изначально поступают из костного мозга незрелыми, в процессе созревания они приобретают ТКР для распознавания антигена и CD4+ или CD8+ рецепторы для согласованной работы с другими иммуноцитами (схема).
Вторичные - периферические органы иммунной системы
Периферические органы иммунной системы - лимфатические узлы, селезенка и лимфоидная ткань, ассоциированная со слизистыми - являются местом встречи антигенов с иммунокомпетентными клетками, местом распознавания антигена и развития специфического ответа, местом взаимодействия иммунокомпетентных клеток, их пролиферации (клональной экспансии), антиген-зависимой дифференцировки и местом накопления продуктов иммунного ответа.
АНТИТЕЛА
Антитела (иммуноглобулины) (АТ, Ig) - в большинстве случаев представлены сывороточными гликопротеинами, входящие в состав фракции гамма-глобулинов. АТ образуют один из основных классов белков крови, составляя 20% массы белка плазмы. Молекула АТ состоит из 2 идентичных тяжелых (Н-цепи) и 2 идентичных легких (L-цепи) аминокислотных цепей, соединенных дисульфидными -S-S- связями (рис.3). В цепях различают вариабельную область (V-область) в N-концевой части и постоянную, или константную, область (С-область). V-область у разных АТ варьирует. V-области L- и Н-цепей образуют Аг-связывающий центр (активный центр АТ, паратоп), или Fab-фрагмент. Константная область молекулы имеет Fc-фрагмент. Аг-связывающий центр, образованный вариабельными областями L- и Н-цепей; именно с ним взаимодействует Аг-детерминанта (эпитоп) антигена. Аг-связывающий центр комплементарен эпитопу Аг по приципу «ключ-замок». Молекула Ig как минимум двухвалентна; такие антитела известны как полные АТ,а мономеры с меньшей валентностью - как неполные АТ. Fc-фрагмент определяет специфичность связывания молекулы Ig с клетками-эффекторами (например, макрофагами, полиморфноядерными лейкоцитами, тучными клетками), несущими на своей поверхности рецепторы Fc-фрагмента.
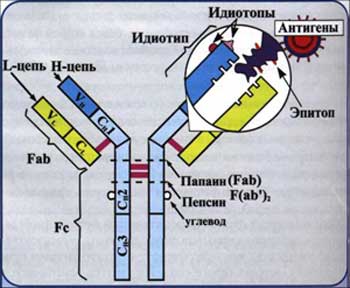
Рис. 6. Структура иммуноглобулина.
Классы иммуноглобулинов и свойства антител
В зависимости от структуры Н-цепей выделяют 5 разных классов (изотипов) АТ: IgA, IgD, IgE, IgG и IgM.
1. IgM -пентамер из 5 субъединиц, соединенных дисульфидными связями, имеет 10 Аг-связывающих участков. IgM - наиболее ранний класс АТ, обнаруживаемый при первичном попадании Аг в организм, т.е. наличие IgM к Аг конкретного возбудителя указывает на наличие острого инфекционного процесса. Молекулы IgM опсонизируют, агглютинируют, преципитируют и лизируют содержащие Аг структуры, а также легко активируют систему комплемента.
2. IgG - Основной класс АТ (до 75% всех Ig), защищающих организм от бактерий, вирусов и токсинов. После первичного контакта с Аг синтез IgM обычно сменяется образованием более дифференцированных IgG. Максимальные титры IgG при первичном ответе наблюдаются на 6-8 сутки. Обнаружение высоких титров IgG к Аг конкретного возбудителя указывает на то, что организм находится на стадии реконвалесценции или конкретное заболевание перенесено недавно.
IgG непосредственно участвуют в реакциях иммунного цитолиза, реакциях нейтрализации, а также усиливают фагоцитоз. Только IgG беременной форсируют плацентарный барьер, проходят черезбазальную мембранутрофобласта в соединительную ткань плода и попадает в капилляры плода. Транспорт IgG через плаценту обеспечивает формирование пассивного иммунитета у плода.
3. I gA (в сывороточном пуле Ig составляют 15-20%) секретируются на поверхность эпителиев, присутствуют в слюне, слезах, молоке, выделяются на поверхность слизистых оболочек, где взаимодействуют с Аг, усиливая защитные свойства слизистых оболочек пищеварительного тракта, дыхательных, половых и мочевыделительных путей. В сыворотке IgA циркулируют в виде мономеров, а в секретируемых SIgA преобладают четырехвалентные димеры, содержащие одну J-цепь и дополнительную полипептидную цепь (синтезируемых эпителиальными клетками секреторный компонент).
4. I gE специфически взаимодействуют с тучными клетками и базофильными лейкоцитами; эти клетки содержат многочисленные гранулы с биологически активными аминами. Защитные потенции IgE направлены преимущественно против гельминтов (нематод).
5. IgD. Биологическая роль этого Ig не установлена; IgD обнаруживают на поверхности развивающихся В-лимфоцитов; в сыворотке здоровых лиц он присутствует в крайне низкой концентрации.
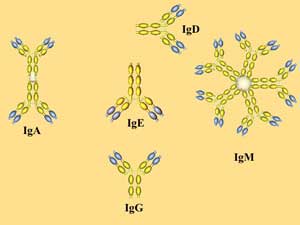
Рис. 7. Антитела разных классов отличаются биологическими свойствами.
Защитные механизмы антител (функции).
АТ помогают предотвратить инфицирование и/или элиминировать возбудитель, активируя при этом несколько механизмов.
- Опсонизация (иммунный фагоцитоз) - связывание АТ с клеточной стенкой микроорганизма с последующим эффективным поглощением образовавшегося комплекса фагоцитом при взаимодействии Fc-фрагмента АТ с соответствующим Fc-рецептором (FcR) на мембране фагоцита.
- Антитоксический эффект. АТ могут связывать и тем самым инактивировать бактериальные токсины.
- Активация комплемента. Антитела (IgM и IgG) после связывания с Аг (микроорганизм, опухолевая клетка и др.) могут активировать каскад сывороточных протеаз (систему комплемента), что приводит к уничтожению индуктора в результате перфорации его клеточной стенки, усилению хемотаксиса и иммунного фагоцитоза.
- Нейтрализация. Взаимодействуя с рецепторами клетки, связывающими бактерии или вирусы, АТ могут препятствовать адгезии и проникновению микроорганизмов в клетки организма-хозяина.
- Антителозависимая цитотоксичность. Распознавание клетки-«мишени» антителами, адсорбированными на Fc-рецепторах NK-клеток, с последующим разрушением их.
Динамика антителообразования
Первичный ответ. Появлению АТ предшествует латентный период продолжительностью 3-5 суток. В это время происходит распознавание Аг и образование клонов плазматических клеток. Затем наступает логарифмическая фаза, соответствующая поступлению АТ в кровь; ее продолжительность - 7-15 суток. Постепенно титры АТ достигают пика и наступает стационарная фаза, продолжительнстью 15-30 суток. Ее сменяет фаза снижения титров АТ, длящаяся 1-6 месяцев.
Вторичный ответ. После антигенной стимуляции часть В- и Т-лимфоцитов циркулирует в виде клеток памяти. Особенности вторичного иммунного ответа - высокая скорость антителообразования, появление максимальных титров АТ и длительное (иногда многолетнее) их циркулирование. Основные характеристики вторичного ответа:
· образование АТ индуцируется значительно меньшими дозами Аг;
· индуктивная фаза сокращается до 5-6 ч;
· среди АТ доминируют IgG с большой аффинностью, пик их наступает раньше (3-5 сут);
· АТ образуются в более высоких титрах и циркулируют в организме длительное время.
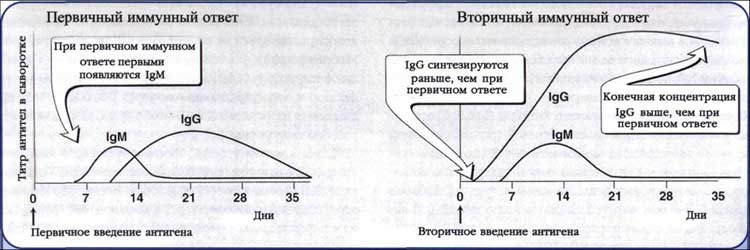
Рис. 8. Антителобразование при первичном и вторичном иммунном ответе.
Тема 23. Формы иммунитета. Виды иммунитета.
Содержание:
- Формы иммунитета
- Виды иммунитета
- Регуляция иммунного ответа

