 2015-05-30
2015-05-30 726
726§ 2. ОПРЕДЕЛЕНИЕ КОНЦЕНТРАЦИИ БАКТЕРИЙ, ФАГОВ, ВИРУСОВ И РИККЕТСИЙ ПО РЕЗУЛЬТАТАМ ПОСЕВА КРАЙНИХ РАЗВЕДЕНИЙ ИССЛЕДУЕМОЙ КУЛЬТУРЫ НА ЖИДКИЕ ПИТАТЕЛЬНЫЕ СРЕДЫ
Для определения концентрации бактерий, фагов, вирусов и риккетсий нередко применяется следующий метод: различные разведения исследуемой культуры высеваются на жидкие питательные среды (для фагов такими средами служат культуры чувствительных бактерий, а для вирусов и риккетсий – культуры чувствительных тканей) и после инкубации отмечаются пробы, в которых максимальные разведения еще вызывают рост определяемого микроорганизма. Доказано, что для возникновения роста достаточно во многих случаях внести единичный микроорганизм. Казалось бы, величина максимального разведения, которое еще вызывает рост, должна быть численно равна количеству живых микроорганизмов в исходной культуре, заключенному в объеме, равному объему посевной дозы. В действительности, однако, интерпретация данных такого рода представляется гораздо более сложной. Дело в том, что при отборе посевной дозы из крайних разведений культуры, содержащих единичные микроорганизмы, вероятность захватить один или несколько микроорганизмов становится соизмеримой с вероятностью вовсе не захватить ни одного микроорганизма. В результате граница между разведениями, посев которых вызывает рост, и разведениями, не вызывающими роста, оказывается обычно весьма размытой, неопределенной: положительные пробы чередуются с отрицательными без какой-либо четкой закономерности. Поэтому, и расчет концентрации исходной культуры, и оценка ошибки этого метода требуют применения своеобразных статистических приемов. Описание одного из наиболее строгих способов приведено в работе Чанга и сотр. [51]. Там же приведена формула Томаса, позволяющая достаточно просто производить необходимые вычисления с точностью, вполне удовлетворяющей требованиям большинства экспериментальных работ:
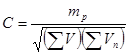 (34).
(34).
Здесь 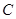 обозначает концентрацию исходной культуры, выраженную в количестве живых микроорганизмов (или каких-то минимальных конгломератов живых микроорганизмов, еще способных вызвать рост в избранных условиях посева и культивирования) на 1 мл;
обозначает концентрацию исходной культуры, выраженную в количестве живых микроорганизмов (или каких-то минимальных конгломератов живых микроорганизмов, еще способных вызвать рост в избранных условиях посева и культивирования) на 1 мл; 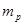 - общее число положительных проб, т.е. таких, в которых наблюдался рост посеянного микроорганизма;
- общее число положительных проб, т.е. таких, в которых наблюдался рост посеянного микроорганизма; 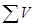 - сумма всех доз, испытанных в данном определении и выраженных в единицах объема (мл) исходной культуры;
- сумма всех доз, испытанных в данном определении и выраженных в единицах объема (мл) исходной культуры; 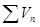 - сумма всех доз, которые дали отрицательный результат (т.е. после посева этих доз не наблюдалось роста). При расчете по приведенной формуле следует принимать во внимание только дозы, посев которых дал хотя бы один отрицательный результат или более; дозы, вызвавшие только положительные реакции, не учитываются.
- сумма всех доз, которые дали отрицательный результат (т.е. после посева этих доз не наблюдалось роста). При расчете по приведенной формуле следует принимать во внимание только дозы, посев которых дал хотя бы один отрицательный результат или более; дозы, вызвавшие только положительные реакции, не учитываются.
Формула Томаса непригодна только в одном случае, если за дозами, которые вызвали 100 % положительных реакций, непосредственно следуют дозы, вовсе не вызвавшие положительных реакций. Например (в порядке уменьшающихся доз) – 5, 5, 5, 0, 0, 0. При этом рекомендуется переписать результаты, уменьшив на единицу число положительных реакций, соответствующих наименьшей из описанных доз, и введя условно одну положительную реакцию для наибольшей из неэффективных доз. В указанном примере это приведет к следующему: 5, 5, 4, 1, 0, 0. После этого можно воспользоваться формулой Томаса. Такая корректировка данных не отражается существенно на точности конечного расчета.
Приведем пример использования формулы (34). Допустим, производится определение концентрации вируса западного энцефаломиелита у лошадей по методу, описанному Брауном [48], т.е. по действию размножающегося вируса на культуру тканей куриного эмбриона, в результате чего смещается рН среды и происходит изменение окраски добавленного индикатора – фенолового красного. В табл. 25 представлены примерные результаты такого определения, произведенного при посредстве заражения культур пятью дозами, взятыми с десятикратным интервалом, причем каждой дозой заражено по пять проб культуры. Прежде чем произвести подстановку представленных в таблице данных в формулу (34), заметим,
Таблица 25
Действие различных доз культуры вируса западного энцефаломиелита лошадей на культуру тканей куриного эмбриона
| № п/п | Доза культуры вируса в мл | Общее число зараженных вирусом проб тканевой культуры | Число проб, в которых происходил рост вируса (изменилась окраска индикатора) | Число проб, в которых роста не отмечено |
| 0,4´10-4 | ||||
| 0,4´10-5 | ||||
| 0,4´10-6 | ||||
| 0,4´10-7 | ||||
| 0,4´10-8 |
что при заражении 1-й и 3-й дозами все пробы оказались положительными. В соответствии с указанным выше правилом результат заражения этими дозами не следует принимать во внимание при расчете. Подставив в формулу результаты заражения 2, 4 и 5-й дозами, получаем:
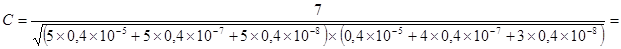
=0,76´106 ед/мл.
Посмотрим теперь, как рассчитывать доверительные интервалы концентраций, находимых с помощью описанного выше метода. Здесь можно обойтись без помощи какой-либо формулы. Для практических целей достаточны, как правило, данные о величинах доверительных интервалов в относительном выражении, приведенные в табл. 26 (по данным работы Чанга и сотр. [51]).
Таблица 26
Коэффициенты для расчета границ доверительных интервалов концентраций, вычисленных по формуле (34)
| Отношение величины каждой предшествующей дозы к последующей* | Число проб среды (тест-культуры), которые заражаются данной дозой | Вероятность, с которой оценивается доверительный интервал | |
| 95 % | 99 % | ||
| 
| ||
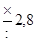
| 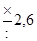
| ||
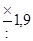
| 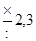
| ||
|
| 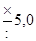
|
*Для каждого определения это отношение должно быть постоянным
51. Chang S.L. and other. – Virology, 1958, 6, 27.
Пояснить применение табл. 26 можно с помощью уже использованного выше примера. Поскольку отношение величины каждой предшествующей дозы к последующей составляет там 10, а число проб на дозу 5, то для вероятности 95 % находим по таблице коэффициент . Следовательно, действительные значения концентрации с вероятностью 95 % заключено в интервале 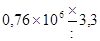 или от
или от 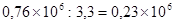 до
до 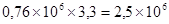 единиц/мл.
единиц/мл.
В заключение раздела отметим, что иногда обработку данных, подобных приведенным выше, производят, вычисляя срединную дозу, вызывающую заражение в среднем 50 % проб, т.е. 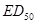 . Это дает не только менее точный результат, но и лишь условную оценку концентрации культуры, в то время как формула (34) позволяет, как правило, вычислить концентрацию, выраженную числом живых микроорганизмов в единице объема.
. Это дает не только менее точный результат, но и лишь условную оценку концентрации культуры, в то время как формула (34) позволяет, как правило, вычислить концентрацию, выраженную числом живых микроорганизмов в единице объема.








